Gist zonder de C-5 sterol desaturase Erg3 vertoont opvallend verhoogde tolerantie voor escin
We veronderstelden dat als escine-toxiciteit wordt gemedieerd door directe interactie met ergosterol, de biosynthesemutanten van ergosterol kunnen verschillen in escine-tolerantie. Van sterolen wordt aangenomen dat ze een sleutelrol spelen bij het handhaven van de homeostase van de plasmamembraandynamiek28echter giststammen zonder enzymen die de laatste vijf stappen in de biosynthese van ergosterol katalyseren (Erg2, Erg3, Erg4, Erg5 of Erg6; Fig. 1b en S1) levensvatbaar zijn. Vanwege de substraatpromiscuïteit van laatstgenoemde enzymen bij de biosynthese van ergosterol, accumuleren deze deletiemutanten een mengsel van sterolstructuren, die van ergosterol verschillen in het aantal en de positie van dubbele bindingen in de sterol B-ring en de sterolzijketen.29. We gebruikten microplaatculturen van de wildtype stam BY4741 (WT) en mutanten uit de Yeast Deletion Collection30, om de groei van de ergosterol-biosynthesemutanten in complex rijk medium (YPD) in aanwezigheid en afwezigheid van escine te beoordelen (Fig. 1C). Sterolextractie en analyse door gaschromatografie-massaspectrometrie bevestigden dat ergosterol zich niet ophoopte in de mutante stammen (Fig. S2 en S3). De minimale remmende concentratie (MIC) van escine voor de WT-stam in YPD-medium was 150 µg/ml. Vergelijkbare MIC's werden waargenomen voor de erg2Δ, erg4Δ, erg5Δ en erg6Δ stammen, maar de groei van de erg3A-stam was ongeremd tot de hoogst geteste concentratie (1000 µg/ml; Fig. S4).
Transcriptoomanalyse
Om de impact van escine op WT-cellen en het mechanisme van escine-tolerantie verder te onderzoeken erg3Δ, we analyseerden de transcriptomen van WT, erg3Δ en erg6Δ-cellen, behandeld met 0 of 100 µg/ml escine in YPD gedurende 1 uur, in schudkolfculturen. De erg6De Δ-stam werd in dit experiment opgenomen, omdat deze stam veel vergelijkbare fenotypes deelt erg3Δ29, maar heeft geen verbeterde escine-tolerantie (MIC 150 µg/ml; Fig. 1C). In het kader van dit experiment hebben we ook de transcriptomen van geanalyseerd erg3Δ en erg6A-cellen die gedurende 75 uur werden behandeld met 1 µg/ml escine.
K betekent clustering van de 2000 genen met de meest variabele expressieniveaus wordt getoond in Fig. S5, met volledige details van de verrijkingsanalyse van de genenset opgenomen in de aanvullende informatie (SI). Cluster B-genen (n = 82) komen over het algemeen op een hoger niveau tot expressie erg3Δ en erg6Δ vergeleken met WT onder alle omstandigheden en, zoals verwacht, is dit cluster verrijkt met genen die verband houden met sterolbiosynthese, steroltransport, siderofoortransport en regulering van transcriptie door glucose. Cluster A-genen (n = 821) worden neerwaarts gereguleerd in WT en erg6Δ als reactie op escine-behandeling. Dit cluster is verrijkt met genen die verband houden met ribosoombiogenese en RNA-verwerking. Cluster C (n = 897) en D (n = 200) genen worden over het algemeen opgereguleerd in WT en erg6Δ als reactie op behandeling met escine, waarbij cluster D-genen in grotere mate opgereguleerd zijn dan cluster C. Cluster D is verrijkt met genen die verband houden met het trehalose-, mannose-, fructose- en glutamaatmetabolisme, evenals met glycolyse en celwandorganisatie. Cluster D is ook verrijkt met genen die geassocieerd zijn met reacties op osmotische, oxidatieve, temperatuur- en hongerstress. Cluster C is verrijkt met genen die verband houden met late nucleofagie, lipidekatabolisme, zwavelassimilatie, de tricarbonzuurcyclus en gluconeogenese.
Differentieel tot expressie gebrachte genen (DEG's) met ≥ 2-voudige differentiële expressie tussen van belang zijnde condities (Fig. 2) worden hieronder in meer detail besproken, met details over de verrijking van de volledige genenset in SI.
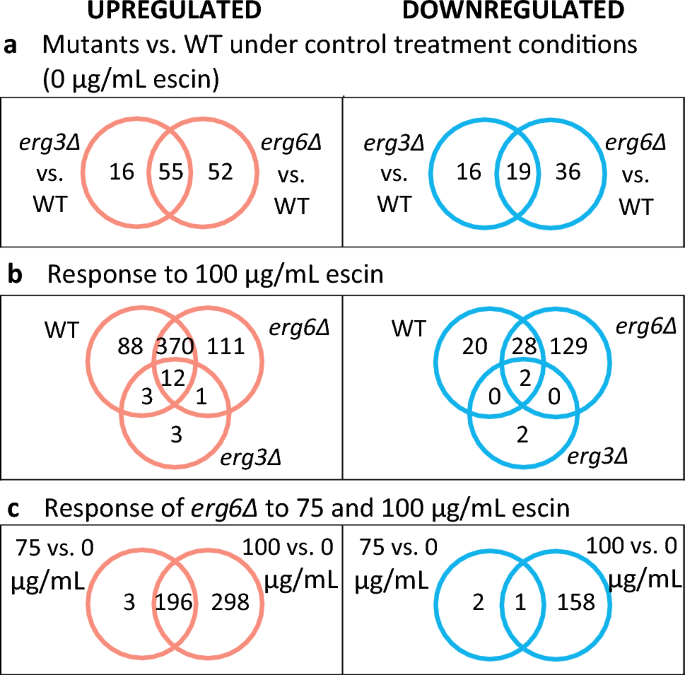
Venn-diagrammen voor differentieel uitgedrukte genen (DEG's). DEG's hebben ≥ 2-voudige differentiële expressie en een False Discovery Rate (FDR) ≤ 0.1.
Impact van ERG3 en ERG6 deletie op het gisttranscriptoom
Het vergelijken van de transcriptomen van stammen onder controleomstandigheden onthult interessante verschillen tussen de mutante stammen en WT. Verwijdering van ERG6 resulteert in een groter aantal DEG's (n = 162) dan ERG3 deletie (n = 106). In beide mutanten zijn opgereguleerde DEG's verrijkt met genen die verband houden met sterolbiosynthese en -transport, inclusief de sterolsensor en transcriptionele activator UPC2 (Fig. S6). Daarentegen wordt slechts een zeer klein aantal genen die betrokken zijn bij de aanvoer van sfingolipiden en fosfolipiden opgereguleerd (YSR3, RSB1, AAF2 en ELO2 in beide stammen, en bovendien SUR1 in erg6een; Afb. S7), hoewel grote veranderingen in de samenstelling van sfingolipiden zijn gerapporteerd voor sterolbiosynthesemutanten31.
Mannoproteïne-genen voor anaerobe respons in de celwand zijn ook sterk opgereguleerd in beide stammen (Fig. S8). Verschillende stappen van de biosynthese van ergosterol vereisen zuurstof en ijzer als cofactoren, en er is sprake van complexe overspraak tussen de regulerende reacties op ergosterol, zuurstof en ijzer.32,33,34,35. Een aantal ijzeruithongeringsreactiegenen36 zijn opgereguleerd erg6Δ (FIT2, FIT3, ARN2, ZIT1, TIS11), maar niet erg3Δ, ten opzichte van WT (Fig. 3A). Onlangs is gemeld dat de ijzergevoelige transcriptiefactor Aft1, die gewoonlijk pendelt tussen het cytoplasma en de kern, zich ophoopt in de vacuolen van ergosterol-verminderde upc2Δ cellen37. Onze gegevens suggereren dat deze signaalroute in verschillende mate kan worden beïnvloed erg6Δ en erg3Δ.
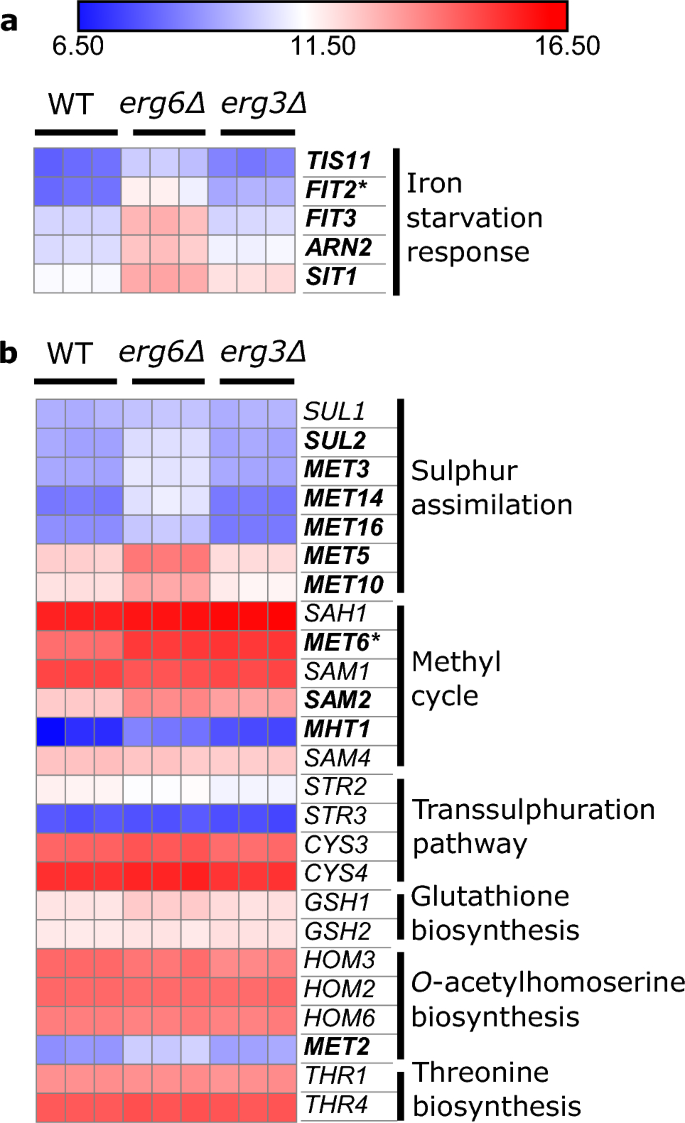
De erg6A-stam vertoont verhoogde expressie van genen voor zwavelassimilatie en genen voor reactie op ijzeruithongering. Expressie van genen met betrekking tot (a), ijzerhomeostase en (b), de methioninebiosyntheseroute, onder controleomstandigheden, eenheden log2(CPM + 4). Vetgedrukt; gen opgereguleerd ≥ 2-voudig in erg6Δ versus WT. Asterisk; gen opgereguleerd ≥ 2-voudig in erg3Δ versus WT (FDR ≤ 0.1).
Methioninemetabolismegenen worden ook opgereguleerd erg6Δ, maar niet erg3Δ (afb. 3B). In het bijzonder zijn de opgereguleerde genen geassocieerd met de zwavel-assimilatieroute (SUL2, MET3, MET14, MET16, MET5, MET10), en de methylcyclus (MET6, SAM2, MHT1) die S-adenosylmethionine (SAM) genereert (Fig. S9). Het Erg6-enzym gebruikt de methyldonor SAM om zymosterol te methyleren38, en een Erg6-Eerder is gebleken dat een tekort aan stam SAM accumuleert, wat werd toegeschreven aan een verminderd SAM-verbruik39.
Beide stammen downreguleren een aantal genen die betrokken zijn bij paring (fig. 4). Dit zijn voornamelijk genen die betrokken zijn bij conjugatie tussen parende schmoo-tips (FUS1, AGA1, AGA2, FIGUUR 1, SHC1), en met name verminderde paringsefficiëntie is eerder beschreven voor ergosterol-biosynthesemutanten40,41.
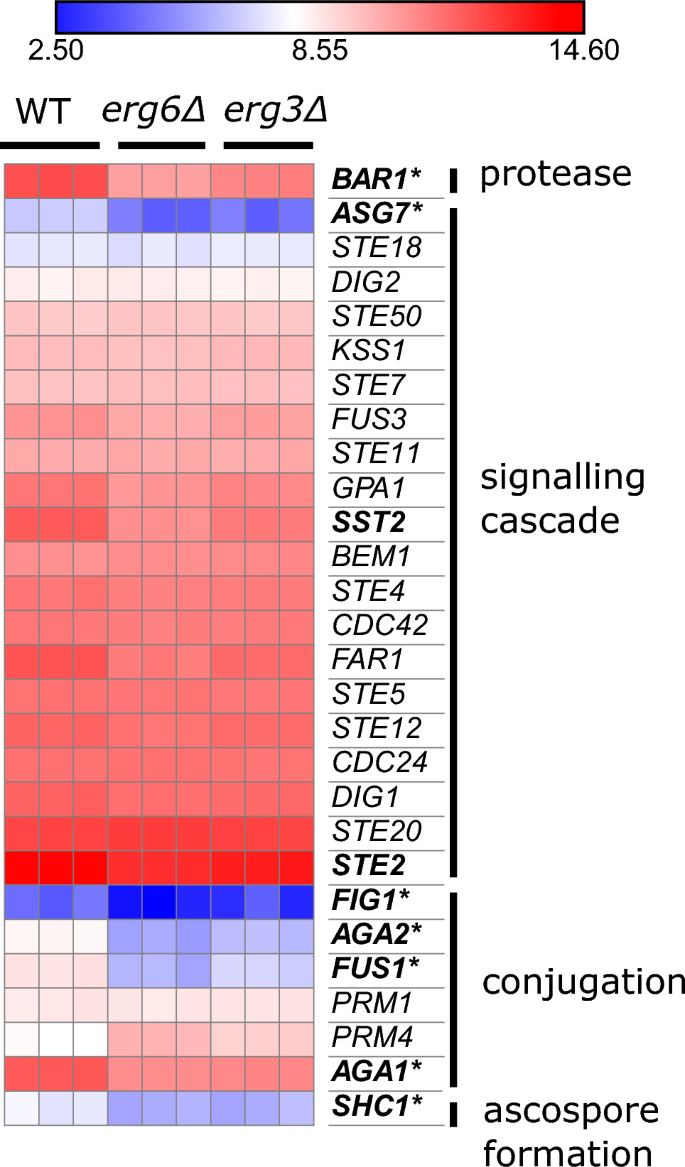
De erg3Δ en erg6A-stammen vertonen verminderde expressie van paringsgenen. Expressie van genen met betrekking tot paring, eenheden log2 (CPM + 4). Vetgedrukt; neerwaarts gereguleerd ≥ 2-voudig in de vergelijking erg6Δ 0 µg/ml escine versus WT 0 µg/ml escine. Asterisk; neerwaarts gereguleerd ≥ 2-voudig in de vergelijkingen erg3Δ 0 µg/ml escine versus WT 0 µg/ml escine (FDR ≤ 0.1).
Beide soorten komen ook tot uiting NVU102 op een lager niveau dan WT (42% van WT-niveau in erg3Δ, en 40% van het WT-niveau in erg6Δ). Hoewel het voornamelijk gelokaliseerd is in de eisosomen van het plasmamembraan, is onlangs gemeld dat Nce102 een rol speelt bij vacuolefusie42, en vacuolen zijn sterk gefragmenteerd in veel ergosterol biosynthese mutante stammen, waaronder erg3Δ, hoewel niet erg6Δ43,44,45.
Het is opmerkelijk dat verschillende DEG's tussen de mutanten en WT een onbekende functie hebben; 20 en 29 opgereguleerde DEG's, en 10 en 13 neerwaarts gereguleerde DEG's, in erg3Δ en erg6Δ respectievelijk.
Transcriptoomreacties op escin-behandeling
Als reactie op 100 µg/ml escin, de WT en erg6A-stammen reguleren respectievelijk 473 en 494 genen (Fig. 2B). Deze DEG's zijn verrijkt met genen die ook geassocieerd zijn met reacties op uitdroging, osmotische stress, oxidatieve stress, temperatuur, chemische behandeling en veranderingen in nutriëntenniveaus (SI). Behandeling van erg6A-cellen met 75 µg/ml escine resulteerden in opregulatie van 199 genen (Fig. 2C). Deze 199°C's zijn verrijkt aan genen die geassocieerd zijn met reacties op oxidatieve stress, chemische stress en uithongering (SI).
De impact van escine-behandeling op genen voor het centrale koolstofmetabolisme wordt getoond in Fig. S10. Als reactie op 100 μg / ml escin, zowel WT als erg6Δ sterk opregulerende genen met betrekking tot de biosynthese van trehalose, glycogeen en glycerol. Aangenomen wordt dat het glucosepolymeer glycogeen in de eerste plaats functioneert als een opslagkoolhydraat, dat weinig invloed heeft op de interne osmotische druk van de cel46,47. Aangenomen wordt dat de disacharide trehalose een sleutelrol speelt bij het beschermen van de structuur van lipidemembranen en eiwitten, door water van lipidedubbellagen en eiwitoppervlakken te verdringen en aggregatie van gedenatureerde eiwitten te voorkomen, wat hun daaropvolgende hervouwing zou voorkomen47. Glycerol fungeert als een belangrijke osmolyt tijdens hyperosmotische stress en vormt de ruggengraat van fosfolipiden en triacylglycerolen voor opslaglipiden48. De verhoging van de biosynthese van trehalose, glycogeen en glycerol maakt deel uit van een algemene reactie op veranderingen in het milieu47,48,49. Bovendien, zowel WT als erg6Δ-stammen reguleren genen die coderen voor eiwitten van het Glucose Induced Degradation Deficient (GID)-complex, dat de gluconeogenese negatief reguleert, ten gunste van glycolyse, door polyubiquitinatie en afbraak van fructose-1,6-bisfosfatase te initiëren50 (Fig. S11).
Als reactie op 100 μg / ml escin, zowel WT als erg6Δ reguleert ook genen die coderen voor enzymen van de γ-aminoboterzuur (GABA) shuntroute (Fig. S12) dat glutamaat afbreekt via GABA. In gist is aangetoond dat deze route belangrijk is voor tolerantie voor hitte en oxidatieve stress51.
De opgereguleerde DEG's zijn ook verrijkt met genen die verband houden met de celwandorganisatie, waaronder sporulatie en biosynthese van chitine (Fig. S13). Overspraak tussen de High Osmolarity Glycerol 1 (Hog1) en Cell Wall Integrity (CWI) mitogeen-geactiveerde proteïnekinase (MAPK) routes betekenen dat het moeilijk is om specifieke spanningen te ontleden die de opregulatie van deze genen initiëren, zonder gebruik te maken van mutanten die defect zijn in specifieke takken van deze signaalroutes52,53,54.
Veel autofagie-gerelateerde genen worden opgereguleerd in WT en erg6Δ als reactie op escine-behandeling (Fig. S14), inclusief kernfagosoomcomponenten, en eiwitten die betrokken zijn bij de cytoplasma-naar-vacuole-route, die selectief hydrolasen aan de vacuole aflevert55. Als reactie op 100 μg/ml escine worden meer autofagie-gerelateerde genen ≥ 2-voudig opgereguleerd in de erg6Δ stam dan WT (11 vs. 7).
De erg6A-stam reguleert ook meer genen dan WT als reactie op 100 μg / ml escine (159 vs. 50), en deze DEG's zijn verrijkt met genen die verband houden met ribosoombiogenese en translatie. Deze processen vergen aanzienlijke hoeveelheden energie56, en hun neerwaartse regulatie maakt deel uit van een algemene stressreactie, waarbij middelen worden omgeleid naar stressresistentie in plaats van celproliferatie57,58.
Als escine ergosterol uit cellen zou afzonderen, of de steroldetectie zou remmen door interactie met ergosterol of een sterolsensor, dan zouden we een transcriptionele verandering in sterolbiosynthesegenen kunnen verwachten, zoals ERG2 en ERG11 als reactie op de escin-behandeling. Deze genen worden gereguleerd door de sterolsensoren en transcriptionele activatoren Upc2 en Ecm22, als reactie op het ergosterolgehalte59,60. Differentiële expressie van genen voor de biosynthese van ergosterol wordt niet waargenomen (Fig. S6), hoewel er sprake is van een statistisch significante, maar kleine opregulering van UPC2 in elke stam (1.3-, 1.9- en 1.5-voudig voor WT, erg6Δ en erg3Δ respectievelijk). Een klein aantal genen gerelateerd aan de toevoer van sfingolipiden wordt opgereguleerd als reactie op escine in WT en erg6Δ (afb. S7), hoewel moet worden opgemerkt dat veel van de regulatie van sfingolipiden posttranslationeel is31.
In tegenstelling tot de grote transcriptoomveranderingen die werden waargenomen voor de WT en erg6Δ spanningen, de erg3Δ-stam reguleert slechts 19 genen in reactie op 100 µg/ml escine, en reguleert 4 naar beneden (Fig. S15). De meeste van deze DEG's worden ook verschillend uitgedrukt in de WT en/of erg6Δ-stam als reactie op escin-behandeling, met uitzondering van HAP4 (codeert een regulator van de ademhaling), YOL163W (beschouwd als niet-functioneel), YGL088W (onbekende functie) en de genen voor mannoproteïne in de celwand DAN1 en DAN4.
De erg3Δ transcriptoom is niet voorbereid op de stress van escinebehandeling
Om te onderzoeken of de escin-respons DEG's in de WT-stam al differentieel worden uitgedrukt in erg3Δ onder controlecondities, mogelijk 'priming' erg3Δ voor de stress van escine-behandeling vergeleken we DEG's in de vergelijkingen WT 100 versus 0 μg / ml escine, en erg3Δ 0 µg/ml escine versus WT 0 µg/ml escine (Fig. S16A). Onder gecontroleerde omstandigheden, de erg3A-stam heeft verhoogde expressie van 18 genen die ook worden opgereguleerd in WT als reactie op escine-behandeling, en lagere expressie van 5 genen die worden opgereguleerd in WT als reactie op escine-behandeling. Een heatmap die de expressieniveaus van deze overlappende genen laat zien, is opgenomen in Fig. S16B. De meeste van deze genen worden ook differentieel gereguleerd in de erg6Δ stam onder controleomstandigheden, en de erg6De A-stam vertoont niet het verbeterde escine-tolerantiefenotype. De uitzonderingen zijn PAI3 (codeert voor een cytoplasmatische proteïnase A-remmer), SRL3 (codeert een celcyclusregulator), HED1 (codeert voor een meiose-specifiek eiwit), MAL31 (codeert een maltosepermease), ARG3 (coderend voor ornithine carbamoyltransferase), COS111 (coderend voor een mitochondriaal eiwit), genen YBR090C en YKR075C van onbekende functie en genen YJR037W en YJL195C waarvan het onwaarschijnlijk is dat ze coderen voor functionele eiwitten.
Over het algemeen laten deze gegevens zien dat escin geen invloed heeft op de groei en een verwaarloosbare invloed op het transcriptoom van erg3Δ cellen. Dit is waarschijnlijk niet te wijten aan erg3Δ-cellen zijn al voorbereid op de spanningen die worden veroorzaakt door de behandeling met escine, en zijn mogelijk toe te schrijven aan de chemische samenstelling van de membraansterolen.
Voormenging van escine met ergosterol, maar niet met brassicasterol, voorkomt door escine gemedieerde groeiremming
Om de hypothese te onderzoeken dat escine toxiciteit medieert door directe interactie met ergosterol, maar niet met de veranderde sterolen die aanwezig zijn in erg3Δ, we hebben de impact beoordeeld van het behandelen van WT-cellen met alleen escine, of escine die vooraf was gemengd met ergosterol in een molaire verhouding van 1: 1. Er werd aangenomen dat als ergosterol een directe interactie aangaat met escine, een mengsel van escine en ergosterol een verminderde impact op de WT-groei zou hebben dan escine alleen, vanwege de verminderde toegankelijkheid van de escine-saponinen. Met name importeren gistcellen geen exogene sterolen onder aërobe omstandigheden37.
Het experiment werd uitgevoerd met behulp van microplaatculturen en volledig supplementmengsel (CSM) media, in tegenstelling tot YPD, aangezien de exacte inhoud van CSM is gedefinieerd, terwijl YPD een rijk medium is dat complexe moleculen van gistextract bevat, die ook kunnen interageren met escine. en/of sterolen. Eerdere studies hebben gemeld dat ergosta-7,22-dienol de overheersende sterol is in gist zonder Erg3, waarbij episterol en ergosta-7-enol zich ook ophopen31,43. Toen het sterolgehalte werd geverifieerd door middel van gaschromatografie-massaspectrometrie (Fig. S2 en S3), de relatieve retentietijd van de grootste sterolpiek in de erg3Δ Total Ion Chromatogram kwam overeen met eerdere rapporten voor ergosta-7,22-dienol, toen de stam werd gekweekt in YPD of CSM.
De MIC van escine in CSM-media werd bepaald op 63 μg / ml voor WT-cellen (Fig. 5B); 2.4 keer lager dan in YPD. Ondertussen groei van de erg3A-stam was ongeremd tot de hoogst geteste concentratie (500 μg/ml). Toen escine vooraf werd gemengd met ergosterol in een molaire verhouding van 1: 1 en het mengsel aan cellen werd toegevoegd, werd de groei van de WT-stam in aanwezigheid van 63 μg / ml escine volledig hersteld (Fig. 5C). Het vooraf mengen van escine met ergosterol herstelde ook het percentage cellen gekleurd met de membraan-ondoordringbare kleurstof propidiumjodide om de behandelingsniveaus onder controle te houden (Fig. S17), wat wijst op verminderde celpermeabiliteit en/of verhoogde cellevensvatbaarheid.
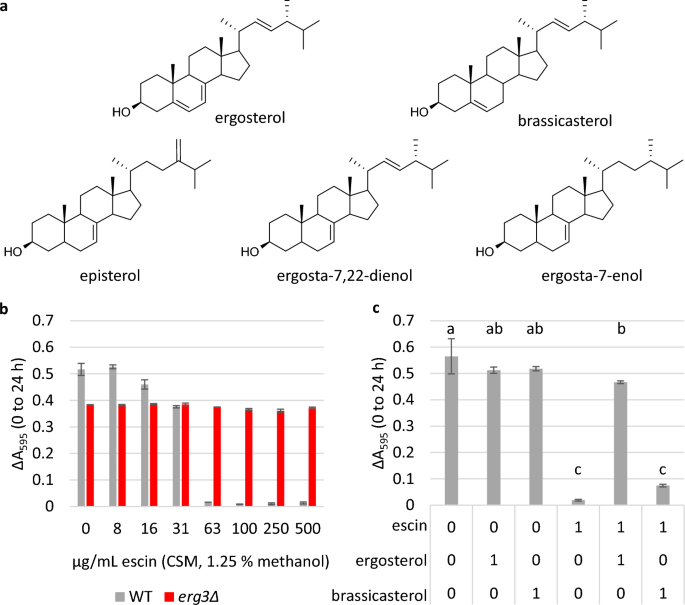
Herstel van de groei wanneer escin wordt voorgemengd met ergosterol, maar niet met brassicasterol. (a) Structuur van ergosterol (de overheersende sterol in S. cerevisiae plasmamembraan), brassicasterol (een plantensterol) en sterolen waarvan is vastgesteld dat ze zich ophopen in het bloed erg3Δ-stam (episterol, ergosta-7,22-dienol, ergosta-7-enol). (b) en (c), Groei in microplaatculturen aangegeven door ΔA595 (0–24 uur). Drie biologische replicatieonderzoeken ± standaarddeviatie (SD). In (b), is de MIC van escin in CSM-media 63 μg/ml voor WT, wat 2.4 keer lager is dan in rijk complex medium. In (c), werden WT-cellen gekweekt in aanwezigheid van 0 of 63 μg/ml escine (1.25% methanol), alleen of vooraf gemengd met ergosterol of brassicasterol in een molaire verhouding van 1:1. Controles met alleen ergosterol en brassicasterol waren opgenomen. Statistieken: one-way ANOVA met post-hoc Tukey HSD-test, condities die niet met dezelfde letter verbonden zijn, zijn significant verschillend (p ≤ 0.05).
De sterolen die aanwezig zijn in erg3Δ zijn van beperkte beschikbaarheid, en daarom hebben we ter vergelijking ook de groei beoordeeld wanneer escin vooraf werd gemengd met het fytosterol brassicasterol, dat ook één dubbele binding in de B-ring heeft, hoewel de positie hiervan tussen C-5 en C- ligt. 6 (afb. 5A). In tegenstelling tot de herstelde groei die werd waargenomen toen escine vooraf werd gemengd met ergosterol, werd herstel van de WT-groei niet waargenomen wanneer escine vooraf werd gemengd met brassicasterol in een molaire verhouding van 1:1 (Fig. 5c).
Dit ondersteunt de hypothese dat escin een directe interactie aangaat met ergosterol en dat kleine veranderingen in de sterolstructuur een grote invloed kunnen hebben op de sterkte van sterol-saponine-interacties.
- Door SEO aangedreven content en PR-distributie. Word vandaag nog versterkt.
- PlatoData.Network Verticale generatieve AI. Versterk jezelf. Toegang hier.
- PlatoAiStream. Web3-intelligentie. Kennis versterkt. Toegang hier.
- PlatoESG. Automotive / EV's, carbon, CleanTech, Energie, Milieu, Zonne, Afvalbeheer. Toegang hier.
- Plato Gezondheid. Intelligentie op het gebied van biotech en klinische proeven. Toegang hier.
- ChartPrime. Verhoog uw handelsspel met ChartPrime. Toegang hier.
- BlockOffsets. Eigendom voor milieucompensatie moderniseren. Toegang hier.
- Bron: https://www.nature.com/articles/s41598-023-40308-0

