ATF3 wordt snel en tijdelijk geïnduceerd tijdens SC vroege activering
Om de transcriptionele regulatoire gebeurtenissen die de overgang van SC-rust naar activering regelen te onderzoeken, hebben we onze eerder gepubliceerde datasets voor transcriptomische profilering opnieuw geanalyseerd36 verkregen van SC's in de afstammingslijn. Kortom, spieren van Pax7-nGFP muizen37 werden in situ gefixeerd door PFA voordat ze werden onderworpen aan het standaard 3 uur lange celdissociatie- en isolatieproces, en FACS-sortering om QSC's te verkrijgen13, of zonder voorafgaande fixatie om vers geïsoleerde vroege activerende SC's (FISC) te verkrijgen (Fig. 1a en aanvullende afb. 1a); FISC's werden 24, 48 of 72 uur in vitro gekweekt om volledig geactiveerde (ASC-24h), prolifererende (ASC-48h) of differentiërende (DSC-72h) cellen te verkrijgen. In overeenstemming met de bevindingen uit andere recente rapporten38,39,40, AP-1-familiegenen inclusief Fos, Atf, jun en Maf subfamilies werden dramatisch geïnduceerd in FISC's in vergelijking met QSC's (Fig. 1b). Opmerkelijk, Atf3 werd scherp geïnduceerd (42.4-voudig) in FISC's maar nam vervolgens snel af bij ACS-24 uur (Fig. 1c, d); dit werd ook bevestigd door RT-qPCR (Fig. 1e), wat aangeeft dat het een snel en tijdelijk reagerend gen is op vroege activering veroorzaakt door de isolatie. Bovendien toonde immunofluorescentie (IF) kleuring de gelijktijdige inductie van ATF3-eiwit in FISC's en een afname van ASC's aan (Fig. 1f). Zelfs op de vers geïsoleerde enkele myofiber waarvan wordt aangenomen dat deze de rustige niche tot op zekere hoogte behoudt en SC-activering minimaliseert41,42, vertoonden de geassocieerde SC's een hoog expressieniveau van ATF3-eiwit onmiddellijk na 2 uur isolatie, dat na 24 uur in kweek afnam (Fig. 1g), wat het idee versterkt dat ATF3 snel wordt geïnduceerd in vroeg geactiveerde SC's. Om de ATF3-inductie in vivo verder te bevestigen, werd BaCl2 geïnjecteerd in de Tibialis Anterior (TA) -spieren van C57BL / 6-muizen om acute spierbeschadiging te veroorzaken. In dit letselmodel36,43,44, weefseldegeneratie met overvloedige infiltraten van immuuncellen wordt normaal gesproken waargenomen op 1 dag na verwonding (dpi); SC's worden snel geactiveerd om uit te zetten als prolifererende myoblasten die vervolgens fuseren om kleine nieuwe myovezels te vormen die embryonale myosine zware keten (eMyHC) tot expressie brengen en worden gekenmerkt door centraal gelokaliseerde kernen (CLN); deze eMyHC+-vezels zijn goed zichtbaar bij 5 dpi. Bij 7 dpi bestaat de spier voornamelijk uit grotere geregenereerde spiervezels met neerwaarts gereguleerde eMyHC-expressie; spierbeschadiging en ontstekingscellen worden grotendeels verwijderd bij 14 dpi, terwijl de geregenereerde spiervezels in omvang blijven groeien en volwassen worden om volledige regeneratie te bereiken rond 30 dpi. Zoals verwacht, werd ATF3-eiwit niet gedetecteerd op niet-verwonde TA-spieren door IF, maar gemakkelijk te zien bij 1 dpi; sommige kleuring fuseerde niet met Pax7 + -cellen, wat in overeenstemming is met de gerapporteerde alomtegenwoordige inductie in meerdere cellen na letsel45. Bij 2 dpi was ATF3-expressie in Pax7 + -cellen sterk verhoogd, gelijktijdig met de volledige activering van SC's in dit stadium (Fig. 1h). Verschillende andere AP-1 familieleden, Atf4, Fos, FosB en juniB, deelde een vergelijkbaar dynamisch expressieprofiel (Fig. 1b-d), wat ook werd bevestigd door RT-qPCR (aanvullend Fig. 1b) en immunofluorescente kleuring (aanvullend Fig. 1c) resultaten. Alles bij elkaar laten onze bevindingen zien dat ATF3 en verschillende andere AP-1-familieleden snel en tijdelijk worden geïnduceerd tijdens vroege SC-activering, wat hun potentiële rol in de regulatie van SC vroege activering suggereert.
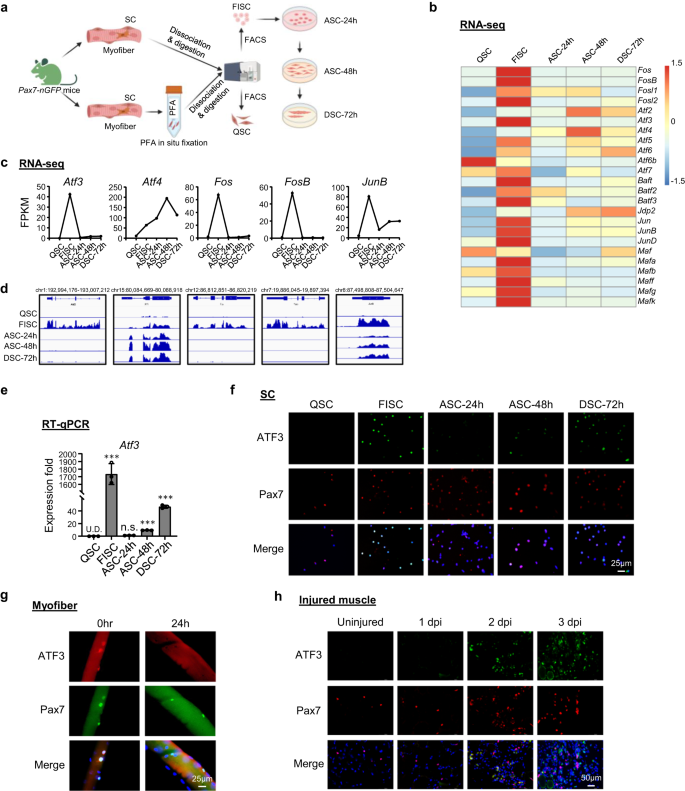
a Schema voor isolatie van rustende satellietcellen (QSC) na in situ fixatie, vers geïsoleerde SC's (FISC) zonder voorafgaande fixatie van spieren van Tg: Pax7-nGFP muizen. FISC's werden vervolgens gekweekt en geactiveerd gedurende 24 (ASC-24 uur), 48 (ASC-48 uur) of 72 uur (DSC-72 uur). RNA's werden geëxtraheerd voor RNA-Seq-analyse. Gemaakt met BioRender.com. b Warmtekaarten die genexpressieniveaus aangeven (Log2 [FPKM]) van AP-1-familie-TF's gedetecteerd door de RNA-Seq. c, d Expressieniveaus (FPKM) en genomische snapshots van Atf3, Atf4, Fos, FosB, juniB mRNA's van de bovenstaande RNA-Seq. e RT-qPCR-detectie van Atf3 in bovenstaande cellen. n = 3 muizen per groep. p = 0.000025, 0.12, 0.0000011 en 0.0000052. f Immunofluorescentie (IF) kleuring van ATF3- en Pax7-eiwitten op de bovenstaande cellen. Schaalbalk: 50 μm. g IF-kleuring van ATF3- en Pax7-eiwitten op enkele myovezels van EDL-spieren onmiddellijk na isolatie of gedurende 24 uur gekweekt. Schaalbalk: 25 μm. h IF-kleuring van ATF3- en Pax7-eiwit op TA-spiersecties van ongedeerde muizen of 1, 2 en 3 dpi. Schaalbalk: 50 μm. Alle staafdiagrammen worden weergegeven als gemiddelde ± SD. Studenten t test (tweezijdig ongepaard) werd gebruikt om de statistische significantie te berekenen (e): *p < 0.05, **p < 0.01, ***p < 0.001. ns geen betekenis. Brongegevens worden geleverd als een brongegevensbestand.
Korte termijn Atf3 deletie versnelt door acuut letsel veroorzaakte spierregeneratie
Om de dissectie van de ATF3-functie in SC's te vergemakkelijken, zijn we de Atf3flox allel46, waarin twee LoxP-sites werden ingevoegd Atf3 exon2, met Pax 7CreERT2/CreERT2-R26RYFP/YFP muis43 controle genereren (Ctrl) (Atf3+ / +– Pax7CreERT2/CreERT2-R26RYFP/YFP) en induceerbare knock-out muizen van Atf3 (Atf3 iKO) (Atf3 flox/flox– Pax7CreERT2/CreERT2-R26RYFP/YFP) (Aanvullende afbeelding. 2a). Na 5 opeenvolgende dagen intraperitoneale (IP) injectie van Tamoxifen (TMX) (fig. 2a), werd succesvolle verwijdering van ATF3 in FISC's bevestigd door WB (aanvullend Fig. 2b) of IF (aanvullend Fig. 2c); de uitputting ervan werd verder bevestigd door IF-kleuring van SC's op enkele myovezels (aanvullend Fig. 2d). Morfologisch vertoonde de 2 maanden oude iKO geen duidelijk verschil met de Ctrl nestgenootmuizen; er werden ook geen veranderingen in lichaamsgrootte en gewicht gedetecteerd (aanvullend Fig. 2e).
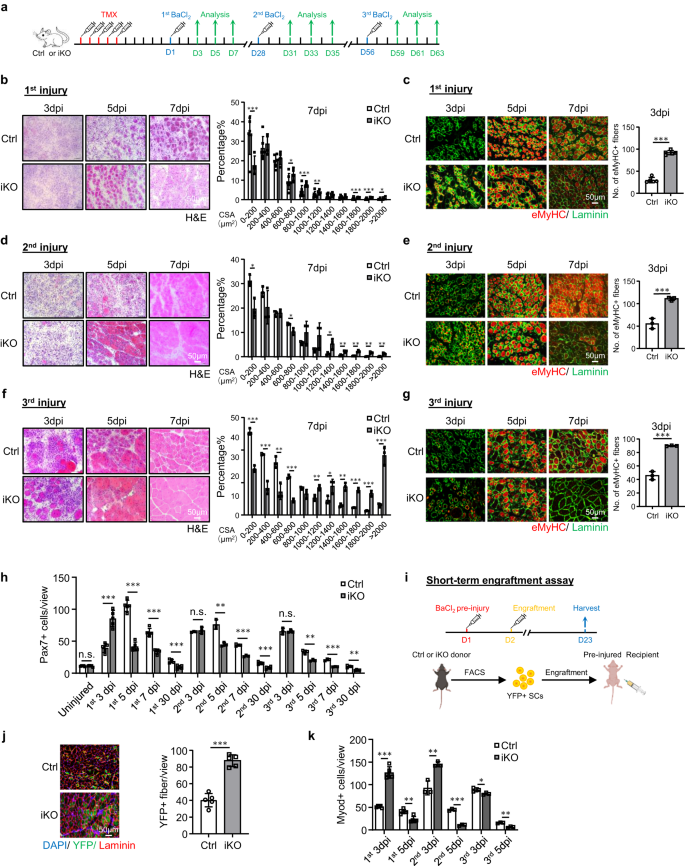
a Schema van het experimentele ontwerp. b H & E-kleuring van TA-spieren verzameld op 3, 5 en 7 dagen na de 1e ronde blessure (dpi). Schaalbalk: 50 μm. Dwarsdoorsnedegebieden (CSA's) van nieuw gevormde vezels werden gekwantificeerd van de TA-spieren bij 7 dpi. p = 0.000066, 0.018, 0.00069, 0.0087, 0.00082, 0.00047 en 0.046. c IF-kleuring van eMyHC (rood) en Laminin (groen) werd uitgevoerd op de bovenstaande TA-spieren. Schaalbalk: 50 μm. p = 0.00000011. d, e H&E en eMyHC-kleuring van de TA-spieren na de 2e ronde blessure. Schaalbalk: 50 μm. p = 0.012, 0.019, 0.033, 0.0057, 0.0022, 0.0022 en 0.0093 (d); p = 0.00021 (e). f, g H&E- en eMyHC-kleuring van de TA-spieren na de 3e ronde blessure. Schaalbalk: 50 μm. p = 0.00015, 0.00087, 0.0063, 0.000074, 0.0089, 0.035, 0.0014, 0.00011, 0.00042 en 0.00071 (f); p = 0.000023 (g). h Pax7+ SC's per weergave werden gekwantificeerd op Ctrl- en iKO TA-spieren op niet-geblesseerde of na de 3 blessurerondes. p = 0.96, 0.00011, 0.0000013, 0.000014, 0.00066, 0.45, 0.0023, 0.00017, 0.00032, 0.95, 0.0022, 0.00019 en 0.0012. i Schema voor engraftment-assay. Gemaakt met BioRender.com. j IF-kleuring van YFP (groen) en Laminin (rood) op de TA-spieren 21 dagen na implantatie. Schaalbalk: 50 μm. n = 5 muizen per groep. p = 0.0000091. k Kwantificering van Myod+ SC's per weergave op Ctrl- en iKO TA-spieren na de 3 blessurerondes. p = 0.0000012, 0.0013, 0.0054, 0.000028, 0.041 en 0.0012. n = 5 muizen per groep gedurende 3, 5, 7 dagen na de 1e verwonding en 30 dagen na de 1e, 2e en 3e verwondings- en implantatietest; n = 3 muizen per groep gedurende 3, 5, 7 dagen na 2e en 3e verwonding. Alle staafdiagrammen worden gepresenteerd als gemiddelde ± SD, student t test (tweezijdig ongepaard) werd gebruikt om de statistische significantie te berekenen (b-h, j, k): *p < 0.05, **p < 0.01, ***p < 0.001. ns geen betekenis. Brongegevens worden geleverd als een brongegevensbestand.
Gezien de snelle inductie van ATF3 tijdens vroege SC-activering, redeneerden we dat het verlies ervan SC-gemedieerde spierregeneratie zou kunnen beïnvloeden. Om dit idee te testen, BaCl2 werd geïnjecteerd in de TA-spieren van Ctrl- of iKO-muizen om 5 dagen na TMX-verwijdering acute spierbeschadiging te veroorzaken (Fig. 2a). Door de bovengenoemde geblesseerde TA-spieren te onderzoeken op 3, 5, 7 en 30 dpi (fig. 2a), vertoonden de iKO-spieren een sterk versnelde regeneratiesnelheid. Door H & E-kleuring (Fig. 2b), bij 5 dpi waren regenererende vezels met CLN's al aanwezig in de iKO maar niet in de Ctrl-spieren. Bij 7 dpi was de beschadigde spier bijna hersteld met homogene vezels en geen teken van immuuncellen in de iKO en de vezelgrootte was ook aanzienlijk groter in vergelijking met de Ctrl (de kwantificering aan de rechterkant). Deze experimentele waarnemingen werden verder ondersteund door IF-kleuring van eMyHC (Fig. 2c); eMyHC+-vezels werden gemakkelijk waargenomen in de iKO, zelfs bij 3 dpi (91.64 in iKO vs. 29.86 in Ctrl), maar namen sterk af met 7 dpi. Consequent was het aantal Pax7+ cellen bij 3 dpi significant hoger in iKO vs. Ctrl maar lager bij 5 dpi en 7 dpi (Fig. 2h en aanvullende afb. 2f). Bovendien hadden de TA-spieren verzameld van de iKO-muizen bij 30 dpi een verhoogd spiergewicht in vergelijking met de Ctrl (43.7 vs. 48.4 mg) (aanvullende figuur XNUMX). 2g) en vergrote spiervezelgrootte (1137.4 vs. 862.8 μm2) (Aanvullende afbeelding. 2h), als gevolg van de verbeterde reparatie.
Verrassend genoeg is het verbeterde regeneratieve vermogen van Atf3 iKO SC's hielden aan na nog een ronde BaCl2-geïnduceerd acuut letsel een maand later gegeven (Fig. 2a). Zoals getoond in Afb. 2d, e, bleef de versnelde regeneratie even sterk op basis van de toegenomen vezelgrootte bij 7 dpi en het aantal eMyHC+-vezels bij 3 dpi na het 2e letsel. Bij 30 dpi vertoonde de iKO-spier ook een verhoogd spiergewicht in vergelijking met de Ctrl (61.7 vs. 55.8 mg) (aanvullende figuur XNUMX). 2i) en licht vergrote spiervezelgrootte (1724.8 vs. 1633.8 μm2) (Aanvullende afbeelding. 2j). Vervolgens werd de 3e blessureronde gegeven (fig. 2a) en de versnelde reparatie was nog steeds duidelijk (afb. 2f, gr), omdat het aantal eMyHC+-vezels bij 3 dpi nog steeds hoger was in de iKO vs. Ctrl-spieren (89.9 vs. 45.7) (Fig. 2g) en een hoger aantal grotere vezels werd waargenomen bij 7 dpi (fig. 2f). Ondanks een licht verhoogd TA-spiergewicht in de iKO in vergelijking met de Ctrl 30 dagen na de 3e blessure (aanvullende afbeelding XNUMX). 2k), was er geen significant verschil in de gemiddelde spiervezelgrootte (aanvullend Fig. 2l), wat suggereert dat de versnelde regeneratie in de iKO-spier uiteindelijk verdween na de 3e blessure.
De engraftment-assay werd vervolgens uitgevoerd om het verbeterde regeneratieve vermogen van de iKO SC's verder te valideren. Zoals geïllustreerd in afb. 2i, 21 dagen nadat YFP + SC's van de Ctrl- of iKO-muizen waren geïnjecteerd in de pre-beschadigde TA-spieren van receptor naakte muizen, werden de TA-spieren verzameld om het regeneratieve vermogen van donorcellen te evalueren op basis van YFP-expressie. We hebben inderdaad een hoger aantal YFP + myofibers waargenomen in de receptormuizen getransplanteerd met de iKO vs. Ctrl SC's (Fig. 2j). Al met al tonen onze resultaten dat aan Atf3 deletie verbetert door acuut letsel veroorzaakte spierregeneratie.
Atf3 deletie veroorzaakt voortijdige SC-activering en pseudo-regeneratie in homeostatische spieren
Om de onderliggende oorzaak voor de verbeterde regeneratieve capaciteit bij ATF3-verlies te ontleden, speculeerden we dat de iKO SC's een snelle activering ondergingen bij BaCl2 blessure. Inderdaad, bij het uitzetten van de dynamiek van MyoD + SC's in de loop van de drie rondes van letsel / regeneratie. een veel groter aantal MyoD+-cellen werd gedetecteerd bij 3 dpi (fig. 2k en aanvullende afb. 2m), maar het aantal cellen daalde snel met 5 dpi. Een dergelijk fenomeen bestond tijdens de eerste twee blessurerondes, maar verdween in de derde ronde (fig. 2k en aanvullende afb. 2m). Vergeleken met de Ctrl, impliceert de snellere activering van iKO-cellen bij acuut letsel de snellere overgang van rust naar activering. Inderdaad, FISC's van de Atf3-iKO-muizen vertoonden een opvallend verhoogde neiging tot het betreden van de celcyclus op basis van EdU-opname 24 uur na kweek (~ 23% toename) (Fig. 3a). Bovendien vertoonden Pax7 + MyoD + -cellen in de iKO een toename van 10% in vergelijking met de Ctrl 24 uur na kweek (Fig. 3b). Bovendien waren de FISC's van de iKO-spieren ook iets groter dan de Ctrl-spieren (86.1 vs. 72.9) (fig. 3c). Consequent hadden geïsoleerde enkele myovezels van de iKO-spieren een significant hogere EdU-opname (14% toename) dan de Ctrl-vezels 36 uur na kweek (Fig. 3d), waardoor de snellere start van de celcyclus van de iKO SC's wordt versterkt. De iKO SC's op vers geïsoleerde enkele myovezels produceerden ook 1.90-voudig en 2.08-voudig meer Pax7 + MyoD + -cellen dan die Ctrl SC's zonder kweken of met 3 uur (Fig. 3e, f). Al met al geven deze resultaten aan dat het verlies van ATF3 resulteert in het vrijkomen van rust en dus snelle SC-activering zonder acuut letsel.
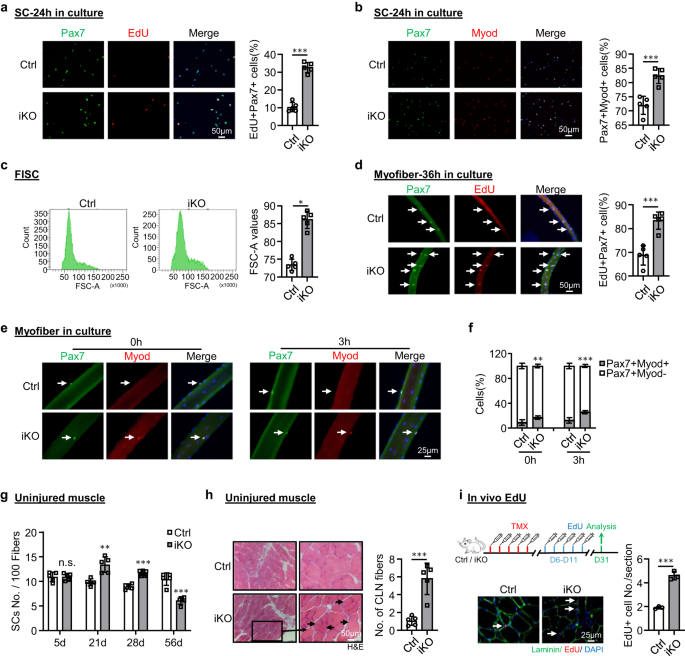
a SC's van Ctrl- of iKO-muizen werden 24 uur gekweekt en 6 uur behandeld met EdU voordat ze werden gekleurd voor EdU (rood) en Pax7 (groen). Schaalbalk: 50 μm. p = 0.0000014. b IF-kleuring van Pax7 (groen) en MyoD (rood) op de bovenstaande SC's gekweekt gedurende 24 uur. Schaalbalk: 50 μm. p = 0.00050. c Representatieve FACS-plots die de grootte van FISC's tonen, gesorteerd op Ctrl- of iKO-spieren. p = 0.050. d Vers geïsoleerde myofibers van Ctrl- of iKO-muizen werden 36 uur gekweekt en 6 uur behandeld met EdU voordat ze werden gekleurd voor EdU (rood) en Pax7 (groen). Schaalbalk: 50 μm. p = 0.00034. e IF kleuring van Pax7 (groen) en MyoD (rood) op de bovenstaande myofibers onmiddellijk na isolatie of gekweekt gedurende 3 uur. Schaalbalk: 25 μm. f Kwantificering van het percentage Myod-Pax7+ en Myod+Pax7+ SC's. p = 0.0098 en 0.00035. g Kwantificering van het aantal Pax7+ SC's per 100 vezels op onbeschadigde Ctrl- of iKO-spieren 5, 21, 28 en 56 dagen na TMX-injectie. p = 0.96, 0.0018, 0.000049 en 0.00018. h H & E-kleuring van de bovengenoemde niet-verwonde spieren 56 dagen na TMX-injectie. Schaalbalk: 50 μm. p = 0.00043. i Boven: Schematisch overzicht van de in vivo EdU-assay uitgevoerd op niet-verwonde Ctrl- of iKO-spier; EdU werd 5 dagen na TMX geïnjecteerd door IP. De spieren werden 21 dagen later verzameld. Lagere IF-kleuring van EdU (rood) en Laminin (groen). Schaalbalk: 25 μm. p = 0.00024. n = 5 muizen per groep (a-h); n = 3 muizen per groep (i). Alle staafdiagrammen worden weergegeven als gemiddelde ± SD, student t test (tweezijdig ongepaard) werd gebruikt om de statistische significantie te berekenen (a-d, f-i): *p < 0.05, **p < 0.01, ***p < 0.001. ns geen betekenis. Brongegevens worden geleverd als een brongegevensbestand.
Het verbeterde regeneratieve vermogen van iKO SC's heeft ons ertoe aangezet om verder te testen of het ATF3-verlies kan leiden tot spontane activering en pseudo-regeneratie in spieren zonder letsel. Vergeleken met de Ctrl was er geen toename van Pax7+ SC's in de iKO TA-spieren op 5 dagen na TMX-injectie, maar een significante toename van Pax7+-cellen in de iKO TA-spieren op 21 (13.4 vs. 9.58) en 28 dagen (11.7 vs. 8.9) (Afb. 3g en S3a), wat wijst op het optreden van spontane activering en expansie van iKO SC's in homeostatische spieren. Interessant is dat 56 dagen na de TMX-injectie het aantal Pax7+-cellen in de iKO-spieren uiteindelijk afnam in vergelijking met de Ctrl (Fig. 3g en S3a); dit ging gepaard met goed zichtbare CLN+ vezels (Fig. 3h), wat wijst op de fusie van de geactiveerde SC-afgeleide myoblasten met bestaande vezels, wat bekend staat als pseudo-regeneratie47. Om deze bevindingen te bevestigen, voerden we vijf doses EdU-injecties uit op 5 dagen na TMX-injectie om aan te tonen dat er meer EdU + -cellen op de spiersecties van de iKO waren dan de Ctrl op 21 dagen na EdU-injectie (Fig. 3i). Evenzo, met een enkele dosis EdU toegediend, werden EdU + -cellen pas 12 uur later gedetecteerd op de geïsoleerde iKO-myovezels of TA-spieren, maar niet op de Ctrl-cellen (aanvullend in Fig. 3b). Al met al suggereren deze resultaten dat Atf3 deletie veroorzaakt de vroegrijpe activering van SC's in homeostatische spieren, dus ATF3 kan functioneren om SC-activering actief te onderdrukken. Dit werd verder versterkt door ATF3 in FISC's tot overexpressie te brengen door een lentivirus (aanvullend Fig. 3c); het remde SC-activering aanzienlijk (Fig. 4a, 20.9% reductie van Pax7+MyoD+ cellen en Fig. 4b, 21.3% reductie van EdU+ cellen). Bovendien werd ATF3 in vivo tot overexpressie gebracht door intramusculaire injectie van de lentivirusdeeltjes bij 1 dpi (Fig. 4c) vertraagde de regeneratie in Ctrl-muizen en stompte ook de versnelde regeneratie in iKO-muizen af (Fig. 4d-h en S3d, bijv).
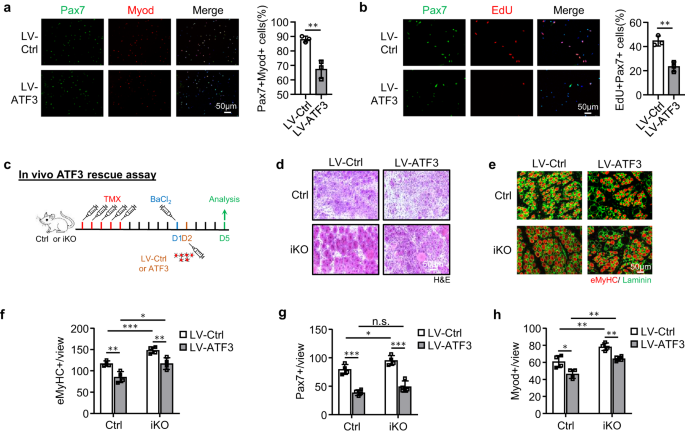
a FISC's van Ctrl- of iKO-muizen werden geïnfecteerd met lentivirussen die ATF3 tot overexpressie brengen en na 7 uur gekleurd voor Pax48 (groen) en Myod (rood). Schaalbalk: 50 μm. p = 0.0050. b EdU-assay werd uitgevoerd en EdU+-cellen werden gekwantificeerd. Schaalbalk: 50 μm. p = 0.0031. c Schema voor overexpressie van ATF3 in vivo. Lentivirussen die ATF3 tot expressie brengen, werden 1 dag na BaCl in Ctrl- of iKO TA-spieren geïnjecteerd2 blessure. De spieren werden 5 dagen na infectie verzameld voor analyse. d H & E-kleuring van de bovenstaande TA-spieren. Schaalbalk: 50 μm. e IF-kleuring van eMyHC (rood) en Laminin (groen) werd uitgevoerd op de bovenstaande TA-spieren. Schaalbalk: 50 μm. f Het aantal eMyHC+-vezels per weergave werd gekwantificeerd. p = 0.0044, 0.00081, 0.015 en 0.0072. g Het aantal Pax7+-cellen per weergave op de bovenstaande TA-spieren werd gekwantificeerd. p = 0.00018, 0.041, 0.098 en 0.00040. h Het aantal Myod+-cellen per weergave op de bovenstaande TA-spieren werd gekwantificeerd. p = 0.016, 0.0058, 0.0010 en 0.0019. n = 3 muizen per groep (a, b); n = 4 muizen per groep (d-h). Alle staafdiagrammen worden weergegeven als gemiddelde ± SD. Studenten t test (tweezijdig ongepaard) werd gebruikt om de statistische significantie te berekenen (a, b, f-h): *p < 0.05, **p < 0.01, ***p < 0.001. ns geen betekenis. Brongegevens worden geleverd als een brongegevensbestand.
Bovendien zagen we dat er een toename van 24% was van MyoD + MyoG + -differentiërende cellen in de geïsoleerde iKO-spiervezels in vergelijking met de Ctrl na 72 uur kweken (aanvullend Fig. 3f), voorstellen dat Atf3 deletie kan de differentiatie van SC-nakomelingen versnellen. Dit werd bevestigd door kleuring voor MyoD en MyoG op SC's die 72 uur waren gekweekt; Een toename van 9% in de dubbele positieve cellen werd gedetecteerd in de iKO vs. Ctrl (aanvullend Fig. 3g). Interessant genoeg was er ook een lichte afname van Pax7 + MyoD-cellen in de iKO-spiervezels in vergelijking met de Ctrl (10.13, 8.48% versus XNUMX, XNUMX%) (aanvullende figuur XNUMX). 3h), wat suggereert dat Atf3-verlies ook kan leiden tot verminderde zelfvernieuwing. Consequent merkten we ook een progressieve afname van het aantal SC's op na elke blessureronde (Fig. 2h en aanvullende afb. 2f).
Lange termijn Atf3 een tekort put de SC-pool uit en belemmert de spierregeneratie
Vervolgens om te testen of het langdurig is Atf3 deficiëntie veroorzaakt SC-uitputting en belemmert spierregeneratie, we veroorzaakten spierbeschadiging door BaCl2 toediening 30 dagen of 120 dagen na verwijdering Atf3 van SC's en onderzocht de geblesseerde TA-spieren op 3, 5 en 7 dpi (Fig. 5a). Dertig dagen na de deletie vertoonde de spierregeneratie (gebaseerd op de spiervezelgrootte en het aantal eMyHC+-vezels) geen significante veranderingen in vergelijking met de Ctrl-muizen (Fig. 5b, c). Het aantal Pax7+ SC was ook vergelijkbaar met 3 en 7 dpi en slechts iets lager in de iKO met 5 dpi (fig. 5d en aanvullende afb. 4a). Daarentegen was 120 dagen na de verwijdering de spierregeneratie duidelijk aangetast in de iKO in vergelijking met de Ctrl-muizen. Een significante toename van kleinere vezels werd waargenomen bij 7 dpi (fig. 5e); het aantal eMyHC+-vezels was veel lager bij 5 dpi maar hoger bij 7 dpi (fig. 5f); het aantal Pax7 + SC's was ook aanzienlijk verminderd bij zowel 3 als 7 dpi (Fig. 5g en aanvullende afb. 4b). Deze resultaten tonen aan dat de lange termijn Atf3 een tekort is schadelijk voor regeneratie door de SC-pool te verkleinen. Interessant is dat wanneer hetzelfde aantal YFP + -cellen geïsoleerd uit de Ctrl- of iKO-muizen 4 maanden na TMX-injectie werd gebruikt in de engraftment-assay (Fig. 5h), werd een hoger aantal YFP+ myofibers waargenomen in de ontvangende naakte muizen getransplanteerd met de iKO-cellen (59.28 vs. 31.72, Fig. 5i), wat suggereert dat het verbeterde regeneratieve vermogen van de iKO-cellen aanhoudt na langdurig ATF3-verlies en dat de verminderde spierregeneratie inderdaad voortkomt uit de verminderde celpool.
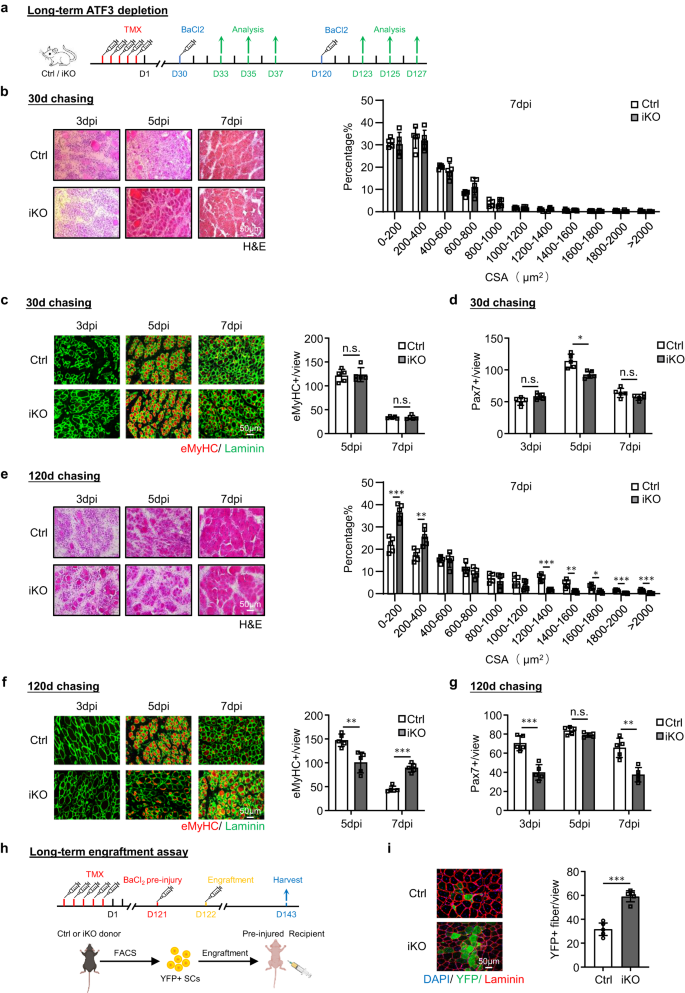
a Schematisch overzicht van de experimentele opzet voor het testen van het effect van lange termijn Atf3 deletie op spierregeneratieproces. 30 of 120 dagen jagen nadat TMX-injectie was gegeven vóór BaCl2 injectie. b Links: H&E-kleuring van de TA-spieren verzameld bij 3, 5 en 7 dpi na de jachtperiode van 30 dagen. Schaalbalk: 50 μm. n = 5 muizen per groep. Rechts: CSA's van nieuw gevormde vezels werden gekwantificeerd van de hierboven gekleurde TA-spier bij 7 dpi en de verdeling wordt weergegeven. n = 5 muizen per groep. c Links: IF-kleuring van eMyHC (rood) en Laminin (groen) werd uitgevoerd op de bovenstaande TA-spieren. Schaalbalk: 50 μm. Rechts: kwantificering van het aantal eMyHC+-vezels per weergave bij 5 en 7 dpi. n = 5 muizen per groep. p = 0.83 en 0.98. d Kwantificering van het aantal Pax7+ SC's per weergave. n = 5 muizen per groep. p = 0.054, 0.0047 en 0.11. e-g De bovenstaande assays/kwantificaties werden uitgevoerd op de Ctrl- of iKO-spieren, verzameld bij 3, 5 en 7 dpi na de jachtperiode van 120 dagen. Schaalbalk: 50 μm. n = 5 muizen per groep. p = 0.00020, 0.0047, 0.000068, 0.0024, 0.013, 0.00019 en 0.00057 (e); p = 0.0034 en 0.000018 (f); p = 0.00031, 0.063 en 0.0010 (g). h Schema voor de engraftment-assay. 120 dagen na TMX-injectie werden FISC's van donormuizen (Ctrl/iKO) geïnjecteerd in naakte muizen van de ontvanger die vooraf gewond waren geraakt. TA-spieren werden 21 dagen na implantatie verzameld voor analyse. Gemaakt met BioRender.com. i Links: IF-kleuring van YFP (groen) en Laminin (rood) op de bovenstaande TA-spieren. Schaalbalk: 50 μm. Rechts: het aantal YFP+-vezels per weergave werd gekwantificeerd. n = 5 muizen per groep. p = 0.000047. Alle staafdiagrammen worden weergegeven als gemiddelde ± SD. Studenten t test (tweezijdig ongepaard) werd gebruikt om de statistische significantie te berekenen (b-g, i): *p < 0.05, **p < 0.01, ***p < 0.001. ns geen betekenis. Brongegevens worden geleverd als een brongegevensbestand.
Om het belang van verder vast te stellen Atf3 bij het opzetten en onderhouden van SC hebben we met succes verwijderd Atf3 in Pax7+ myogene voorlopers door de Atf3flox allel met een niet-induceerbaar Pax 7Cre-R26RYFP transgene muizenstam48, waarin het Cre-recombinase al in E7 tot expressie wordt gebracht in Pax9.5 + -voorlopercellen (aanvullend in Fig. 5a-c). Interessant is dat de cKO-spieren 84% toename van Pax7 + SC's vertoonden in vergelijking met de Ctrl ongeveer 1 maand na de geboorte toen de SC-pool werd opgericht49, maar 79% afname bij volwassen muizen van twee maanden oud (aanvullend Fig. 5e), wat aangeeft dat ATF3 vereist is voor SC-onderhoud op volwassen leeftijd, maar overbodig is voor SC-vestiging. Een verlaagd TA-spiergewicht werd gedetecteerd in cKO op een leeftijd van 1 maand (29.42 mg versus 34.88 mg, aanvullende figuur XNUMX). 5f) maar geen significant verschil in lichaamsgewicht (12.66 mg vs. 12.42 mg, aanvullende figuur XNUMX). 5d) en vezelgrootte (861.96 μm2 versus 848.71 μm2Aanvullende Fig. 5g), is het dus moeilijk te concluderen of spierhypertrofie optreedt bij de jonge cKO-muizen. Naar verwachting vertoonden cKO-cellen, toen de volwassen SC's werden geïsoleerd en gekweekt, een veel hogere neiging tot activering, beoordeeld door zowel Pax7 + Myod + -kleuring (aanvullend Fig. 5h, 23.8% toename in cKO vs. Ctrl) en EdU-assay (aanvullend Fig. 5i, 99.9% stijging). Dienovereenkomstig was het regeneratieve vermogen na acuut letsel duidelijk aangetast in cKO-spieren (aanvullend Fig. 5j-l). Daarom veroorzaakt het verlies van ATF3 vroegtijdige SC-activering en leidt dit tot de uiteindelijke vermindering van de SC-pool.
Atf3 deletie verbetert SC-activering tijdens vrijwillige en uithoudingsoefeningen
Kortdurende en niet-inspannende vrijwillige inspanning (VE) veroorzaakt geen SC-activering, maar duurtraining (EE) activeert SC's50,51,52,53,54. Om de functie van ATF3 bij het voorkomen van SC-activering verder te bewijzen, hebben we geprobeerd te bepalen of de ATF3-uitputting de door VE geïnduceerde SC-activering beïnvloedt. Onmiddellijk na vijf doses TMX-injectie werden Ctrl- en iKO-muizen onderworpen aan een vastgesteld vrijwillig wielloopregime; de muizen kregen toegang tot vrij draaiende loopwielen; een stabiele VE-routine werd bereikt binnen 7 dagen training en werd voortgezet gedurende nog eens 21 dagen lang VE (fig. 6a). De dagelijkse loopafstand werd geregistreerd en er werd geen significant verschil gedetecteerd tussen Ctrl (14.38 km) vs. iKO (13.78 km) muizen (aanvullend Fig. 6a, b).
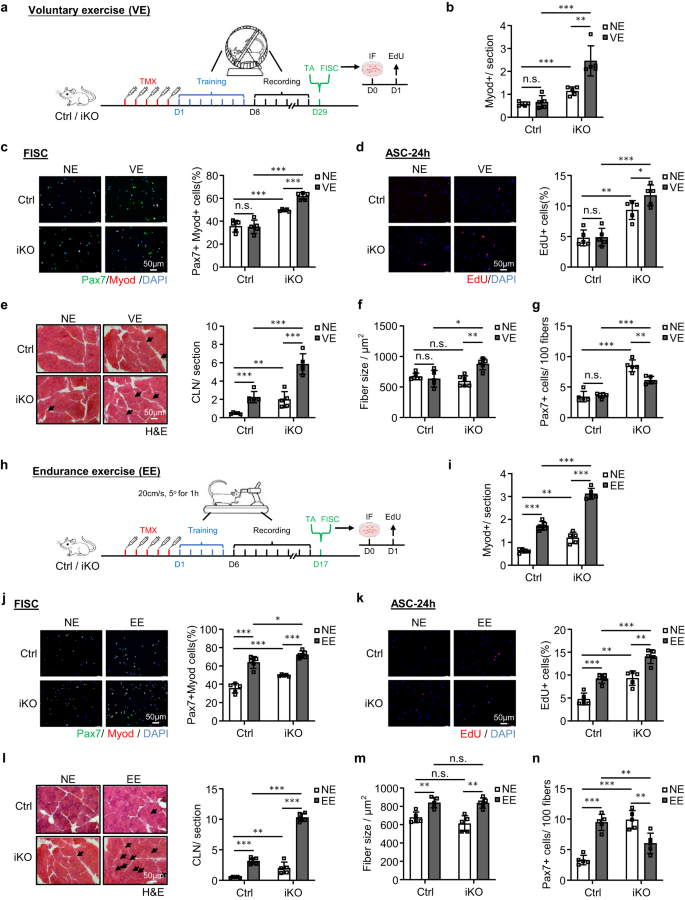
a Schema van het experimentele ontwerp voor VE. b Kwantificering van Myod+-cellen op de bovenstaande TA-spieren. p = 0.54, 0.00024, 0.00049 en 0.0024. c IF-kleuring van Pax7 (groen) en MyoD (rood) op SC's geïsoleerd uit de bovenstaande muizen en 24 uur gesorteerd gekweekt. Schaalbalk: 50 μm. p = 0.91, 0.00035, 0.000018 en 0.000032. d EdU (rood) werd gekleurd op SC's gesorteerd van de bovenstaande muizen en 24 uur gekweekt. Schaalbalk: 50 μm. p = 0.90, 0.0010, 0.00013 en 0.046. e H & E-kleuring van de bovenstaande TA-spieren. Schaalbalk: 50 μm. p = 0.00043, 0.0073, 0.00021 en 0.00047. f Vezelgrootte van de bovenstaande TA-spieren (e). p = 0.54, 0.19, 0.012 en 0.0017. g Kwantificering van Pax7 + SC's op de bovenstaande TA-spieren. p = 0.66, 0.000023, 0.000052 en 0.0017. h Schema van het experimentele ontwerp (EE). i Kwantificering van Myod+-cellen op de TA-spieren verzameld van de bovenstaande Ctrl- en iKO-muizen met of zonder EE. p = 0.0000026, 0.0010, 0.0000073 en 0.0000015. j IF-kleuring van Pax7 (groen) en MyoD (rood) op SC's gesorteerd van de bovenstaande muizen en 24 uur gekweekt. Schaalbalk: 50 μm. p = 0.000062, 0.00035, 0.035 en 0.0000018. k EdU (rood) werd gekleurd op SC's geïsoleerd uit de bovenstaande muizen en 24 uur gekweekt. Schaalbalk: 50 μm. p = 0.00031, 0.0010, 0.00034 en 0.0012. l H & E-kleuring van de bovenstaande TA-spieren. Schaalbalk: 50 μm. p = 0.0000039, 0.0073, 0.000000071 en 0.00000026. m Vezelgrootte van de bovenstaande TA-spieren (l). p = 0.0022, 0.19, 0.91 en 0.0012. n Kwantificering van Pax7 + SC's per op de bovenstaande TA-spieren. p = 0.000018, 0.000024, 0.0067 en 0.0049. n = 5 muizen per groep (b-g, i-n). Alle staafdiagrammen worden weergegeven als gemiddelde ± SD. Studenten t test (tweezijdig ongepaard) werd gebruikt om de statistische significantie te berekenen (b-g, i-n): *p < 0.05, **p < 0.01, ***p < 0.001. ns geen betekenis. Brongegevens worden geleverd als een brongegevensbestand.
Naar verwachting werden de uitdrukkingen van AP-1-familieleden geïnduceerd door de VE in Ctrl SC's (aanvullend in Fig. 6c). Komt ook overeen met eerdere rapporten54, induceerde de VE geen SC-activering in de Ctrl-muizen omdat er geen MyoD + -cellen werden gedetecteerd op de spiersecties voor en na de VE (Fig. 6b en aanvullende afb. 6d); consistent, Pax7 + MyoD + kleuring van FISC's (Fig. 6c) of EdU-kleuring van ASC's gekweekt gedurende 24 uur (Fig. 6d) onthulde geen verhoogd activeringspercentage voor en na de VE. Daarentegen werden de iKO-cellen gemakkelijk geactiveerd door de VE (Fig. 6b-d en aanvullende afb. 6d). Consistent ontdekten we in de iKO-spieren ook een significant verhoogd aantal CLN-vezels (toename van 136%) (Fig. 6e) en duidelijke hypertrofie (vezelgrootte toegenomen met 43.3%) na de VE (Fig. 6f). In de Ctrl-spieren was het aantal CLN verhoogd na de VE, maar niet genoeg om hypertrofische groei te veroorzaken (fig. 6e, f). De bovenstaande bevindingen versterken dus het idee dat ATF3-verlies snelle activering van SC's door de VE veroorzaakt. Als gevolg hiervan was er een verminderde SC-pool (38.2% afname van Pax7+ FISC's) aan het einde van het VE-regime (Fig. 6g en aanvullende afb. 6e).
Omdat bekend is dat EE SC-activering en spierhypertrofie veroorzaakt50,51,52,53, hebben we vervolgens onderzocht of de Atf3 deficiëntie beïnvloedt de door EE geïnduceerde SC-activering. Daartoe werden Ctrl- en iKO-muizen onderworpen aan een loopbandregime55,56 waarbij de loopband gedurende 5 minuten op een helling van 20° en een snelheid van 60 cm/s werd gezet. Na vijf doses TMX-injectie werden de muizen getraind gedurende een aanpassingsperiode van 5 dagen, gevolgd door een duurtraining van 10 dagen (+EE) of, als controleconditie, zonder enige training (NE) (Fig. 6h). Zoals verwacht, werden de expressies van AP-1-familieleden snel geïnduceerd door de EE in SC's (aanvullend Fig. 6c). Bovendien veroorzaakte de EE duidelijke SC-activering in de Ctrl-spieren, maar de activatie was veel sterker in de iKO-spieren (Fig. 6i-k en aanvullende afb. 6f). Consequent werden CLN-vezels geïnduceerd in zowel Ctrl- als iKO-spieren na de EE; en een hogere toename van CLN-vezels werd gezien in de iKO in vergelijking met de Ctrl-spieren (Fig. 6l). Bovendien vertoonden zowel Ctrl- als iKO-spieren hypertrofische groei na de EE, maar er werd geen verschil gedetecteerd (Fig. 6m). Aan het einde van het EE-regime was het aantal Pax7+ FISC's aanzienlijk toegenomen in de Ctrl maar afgenomen in de iKO (Fig. 6n en aanvullende afb. 6g), wat wijst op de snelle uitputting van de iKO SC-pool. Al met al geven deze bevindingen aan dat het verlies van ATF3 SC-activering door VE veroorzaakt en ook SC-activering door EE verbetert.
ATF3 reguleert H2B genexpressie en nucleosoompatronen
Al met al suggereren de bovenstaande resultaten een essentiële rol van ATF3-inductie bij het voorkomen van vroegrijpe SC-activering, om het onderliggende moleculaire mechanisme te verlichten, hebben we de ATF3-transcriptie-output geprofileerd door RNA-Seq uit te voeren in FISC's verzameld uit de Ctrl- en iKO-spieren (Fig. 7a). Een totaal van 1866 transcripten (rode stippen) (79.9%) waren naar boven gereguleerd in de iKO in vergelijking met de Ctrl, terwijl 469 (blauwe stippen) (20.1%) naar beneden waren gereguleerd (Fig. 7bAanvullende Fig. 7a en aanvullende gegevens 1), wat suggereert dat het ATF3-verlies wereldwijde transcriptionele activering induceerde. De GO-analyse onthulde dat de opwaarts gereguleerde genen waren verrijkt voor "mitochondrion", "extracellulaire matrix", enz. (Fig. 7c en aanvullende gegevens 1). Merk op dat verhoogde mitochondriale activiteit en extracellulaire matrixexpressie eerder zijn beschreven in de GAlarm cellen10, wat suggereert dat ATF3 functioneert om de expressie van deze activerende genen te onderdrukken, waardoor voortijdige SC-activering wordt voorkomen. De naar beneden gereguleerde genen waren interessant genoeg verrijkt voor "nucleosoom" en "nucleosoomassemblage", enz. (Fig. 7d en aanvullende gegevens 1). Met name de genen die coderen voor Histone-eiwitten waren sterk vertegenwoordigd onder de naar beneden gereguleerde genen (Fig. 7e), wat ook werd bevestigd door RT-qPCR (19-46% afname) (aanvullend Fig. 7b). Voor histon coderende genen zijn typisch georganiseerd in clusters met meerdere genen en het H2B-eiwit wordt gecodeerd door 2 genclusters waarvan 15 op Chr13 een Hist1h2b cluster en 2 op Chr3 vormen een Hist2h2b cluster (aanvullend Fig. 7c). Opvallend genoeg 13 van de 17 H2b genen waren neerwaarts gereguleerd in de iKO SC's (Fig. 7e). Daarnaast hebben we ook RNA-Seq uitgevoerd op FISC's geïsoleerd uit de Ctrl en Atf3-cKO-muizen en vergelijkbare resultaten werden verkregen (aanvullend Fig. 7d-g en aanvullende gegevens 3). Deze resultaten tonen aan dat het ATF3-verlies afneemt H2b genexpressie.
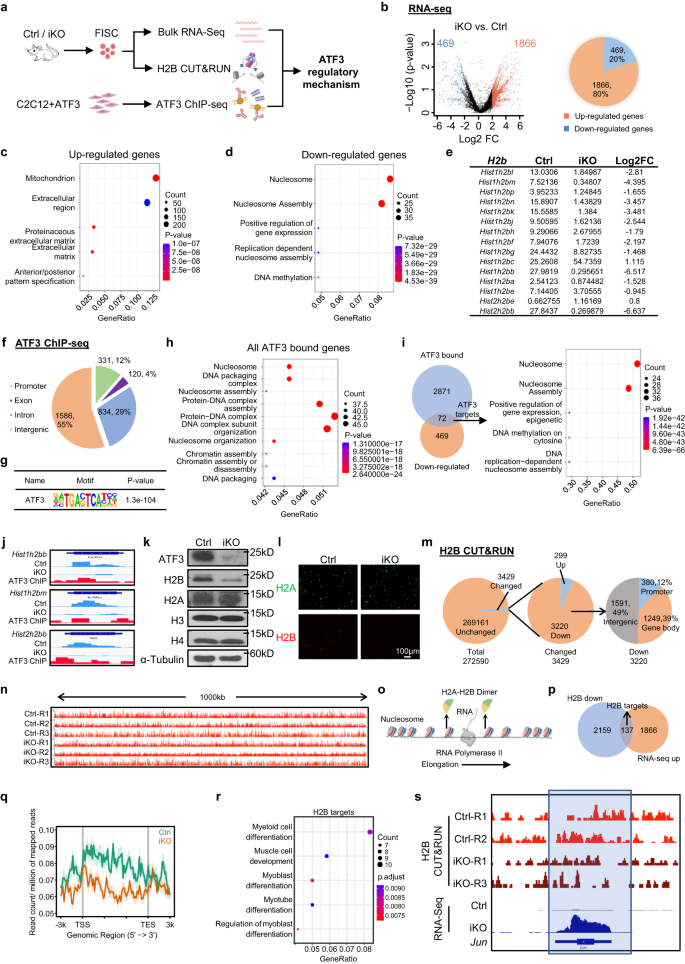
a Schema van het experimentele ontwerp. Gemaakt met BioRender.com. b DEG's werden geïdentificeerd uit de bovenstaande Ctrl vs. iKO RNA-seq. c, d GO-analyse van de bovenstaande op en neer gereguleerde genen. e FPKM en Log2FC van H2b genen in iKO vs. Ctrl. f Genomische verdeling van 2871 ATF3-bindingspieken. g Verrijking van canonieke ATF3-motieven in de bovenstaande bindingsgebieden. h GO-analyse voor alle bovenstaande genen met ATF3-binding. i Links: Venn-diagrammen die de overlapping (72 genen) tonen tussen het hierboven geïdentificeerde ChIP-Seq-doelwit (2871) en de naar beneden gereguleerde genen (469). Rechts: GO-analyse van de bovenstaande 72 genen onthulde een extreme verrijking van histon-gerelateerde termen. j Genomische momentopnamen van 3 van de hierboven geïdentificeerde H2b genen met ATF3-binding in hun TSS's (ChIP-Seq-tracks) en neerwaarts gereguleerd door ATF3-deletie (RNA-Seq-tracks). k ATF3-, H2A-, H2B-, H3- en H4-eiwitten werden gemeten met Western-blot in FISC van Ctrl- en iKO-muizen. α-Tubuline werd gebruikt als laadcontrole. l IF-kleuring van H2A (groen) en H2B (rood) op de bovenstaande FISC's. Schaalbalk: 100 μm; n = 3 muizen per groep. m Cirkeldiagrammen met het aantal bakken met H2B gewijzigd (3429, 299 omhoog en 3220 omlaag) en ongewijzigd (269161) (links en midden), en de genomische verdeling van omlaag gereguleerde H2B-signalen (rechts). n Genomic snapshots van een 1000 kb regio Integrative Genomics Viewer (IGV) track van H2B-signaal in Ctrl vs. iKO op Chr4. o Schema van het RNA-polymerase II-verlengingsproces tijdens DNA-transcriptie. Gemaakt met BioRender.com. p Venn-diagrammen die de overlapping (137 genen) tonen tussen de bovenstaande regio's met neerwaarts gereguleerde H2B CUT&RUN-signalen (2159) en de opwaarts gereguleerde genen (1866) van de bovenstaande (A) RNA-Seq. q Metaplots tonen gemiddelde H2B CUT&RUN-signalen 3 kb stroomopwaarts tot stroomafwaarts van TSS. r GO-analyse van de bovenstaande 252 genen van (P). s Genomische snapshots van Jun gen met neerwaarts gereguleerd H2B CUT&RUN-signaal en opwaarts gereguleerd RNA-Seq-signaal in iKO vs. Ctrl. Regio's met H2B-signaalreductie in iKO vs. Ctrl zijn gemarkeerd in een blauw kader. Brongegevens worden geleverd als een brongegevensbestand.
Vervolgens hebben we ChIP-Seq uitgevoerd op de C2C12-myoblasten met exogene ATF3-overexpressie als een surrogaatsysteem om directe bindingsdoelen van ATF3 te definiëren (Fig. 7a en aanvullende afb. 7h). In totaal werden 2871 bindingspieken geïdentificeerd in 869 genen, waarvan 60%, 28% en 9% zich in respectievelijk intergene regio's, introns en promoters bevinden (Fig. 7f en aanvullende gegevens 2). Deze pieken waren verrijkt voor het bekende ATF3-bindingsmotief, TGACTCA (Fig. 7g), wat wijst op de goede datakwaliteit. GO-analyse van deze ATF3-gebonden genen duidde op opmerkelijke verrijking voor de termen gerelateerd aan nucleosoom (Fig. 7h en aanvullende gegevens 2). Door de RNA-Seq- en ChIP-Seq-resultaten te integreren, bevatten 72 neerwaarts gereguleerde genen ATF3-bindingsplaatsen en opnieuw werden ze verrijkt voor nucleosoomgerelateerde GO-termen en vele waren histon-genen (Fig. 7i en aanvullende gegevens 2). Met name 8 van de 13 down-gereguleerd H2b genen hadden ATF3-bindingsplaatsen in hun promoters (Fig. 7i, jAanvullende Fig. 7i en aanvullende gegevens 2). Ondertussen bezaten 112 opwaarts gereguleerde genen ATF3-binding en waren verrijkt voor mitochondrion-gerelateerde GO-termen (aanvullend Fig. 7j en aanvullende gegevens 2). De Western-blot (fig. 7k) en IF-kleuring (Fig. 7l) resultaten bevestigden een substantiële vermindering of bijna verlies van H2B-eiwit in de iKO FISC's, terwijl andere histon-eiwitten zoals H3, H4 en H2A ongewijzigd bleven. Deze resultaten tonen dat aan H2b genen zijn bonafide transcriptiedoelen van ATF3.
Wereldwijde vermindering van histonen leidt tot een verminderde hoeveelheid nucleosomen, een grotere afstand tussen de nucleosomen en verandert de bezetting van de nucleosomen, waardoor de transcriptie wordt beïnvloed57,58,59, om te bepalen of het H2B-verlies in de iKO SC's de positionering en bezetting van het nucleosoom verandert, CUT&RUN-assay60,61 werd uitgevoerd om de genomische lokalisatie van H2B in Ctrl- en iKO-cellen in kaart te brengen (Fig. 7a). In totaal werden 272590 bins genoombreed gedefinieerd uit de drie replica's (aanvullende gegevens 4). Onverwacht waren de gemiddelde H2B CUT&RUN-signalen grotendeels ongewijzigd in de iKO vs. Ctrl met slechts 3429 bins (1.4%) die veranderingen vertoonden (Fig. 7m, n). Desalniettemin vertoonden 3220 van de gewijzigde bakken inderdaad een verminderde H2B-verrijking in verschillende genomische regio's, waaronder promotor (12%), genlichaam (39%) en intergene regio's (49%) (Fig. 7m). Omdat het nucleosoom gedeeltelijk uiteenvalt door de verwijdering van één H2A-H2B-dimeer om Pol II-transcriptie te vergemakkelijken61 (Fig. 7o), veronderstelden we dat het lokale verlies van H2B de destabilisatie van het nucleosoom zou kunnen vergemakkelijken om de transcriptie van geassocieerde genen te bevorderen. Inderdaad, door RNA-Seq-gegevens te kruisen, vertoonden 137 opwaarts gereguleerde genen verminderde H2B-signalen op hun promoters of genlichamen (Fig. 7p, q en aanvullende gegevens 4). Deze H2B-doelgenen waren verrijkt voor GO-termen gerelateerd aan spierdifferentiatie en ook opgenomen ID3, IGF-1 en Jun, enz. (Afb. 7r, s), wat consistent was met de activeringsstatus van de iKO SC's11,62,63. Alles bij elkaar laten deze bevindingen zien dat het verlaagde H2B-niveau de vroegrijpe SC-activering na het ATF3-verlies zou kunnen mediëren.
H2B bemiddelt de ATF3-functie bij SC-activering en spierregeneratie
Om de functionele link tussen ATF3 en H2B bij SC-activering en spierregeneratie te verstevigen, ontdekten we dat het tot overexpressie brengen van H2B door transfectie van een pcDNA-H2B-plasmide in FISC's inderdaad de versnelde activering van Atf3 iKO-cellen onderdrukte, maar geen impact op Ctrl-cellen (Fig. 8a, b). En in vivo hebben we ook H2B tot overexpressie gebracht (aanvullend Fig. 7k) door intramusculaire injectie van lentivirale deeltjes bij 1 dpi (fig. 8a). Zoals getoond in Afb. 8c-g, herstelde dit ook de door acute verwonding geïnduceerde regeneratie in de Atf3-iKO-muizen: een significant lager aantal eMyHC + -vezels werd waargenomen 5 dpi (Fig. 8d, e), wat gepaard ging met een verminderd aantal Pax7 + -cellen (Fig. 8f) en MyoD+ cellen (Fig. 8g). Al met al valideren deze bevindingen dat H2B-verlies de vroegrijpe SC-activering en verbeterde spierregeneratie medieert die worden waargenomen bij ATF3-verlies.
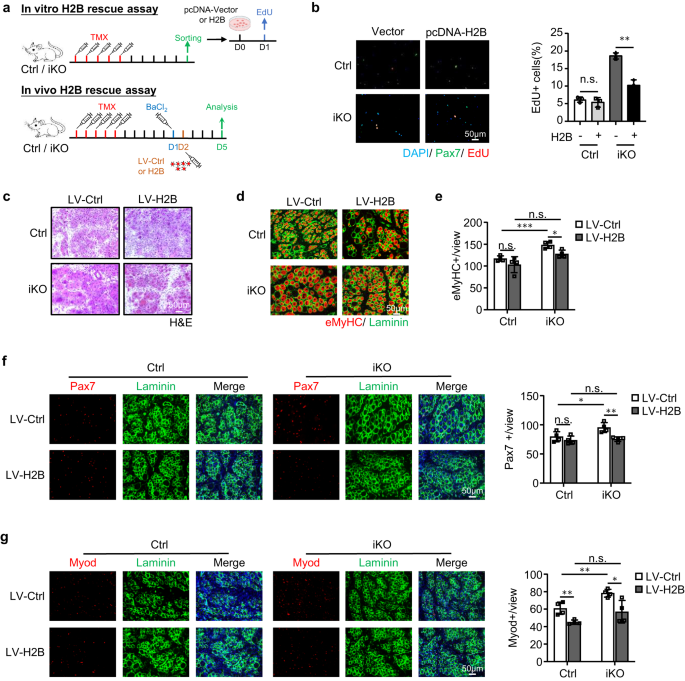
a Boven: Schema voor overexpressie van H2B in vitro. FISC's van Ctrl- of iKO-muizen werden getransfecteerd met een controle (Ctrl) of pCDNA-H2B-plasmide en er werd een EdU-assay uitgevoerd om SC-activering te beoordelen. Onder: Schema voor overexpressie van H2B in vivo via lentivirus. H2B dat lentivirus tot expressie brengt, werd 1 dag na BaCl in Ctrl- of iKO TA-spieren geïnjecteerd2 letsel en de spieren werden 5 dagen na infectie verzameld voor analyse. b Links: de hierboven getransfecteerde cellen werden 24 uur gekweekt voordat ze 6 uur met EdU werden behandeld; EdU-positieve cellen werden gekleurd en gekwantificeerd. Schaalbalk: 50 μm; n = 3 muizen per groep. p = 0.44 en 0.0013. c H & E-kleuring van de bovenstaande TA-spieren verzameld bij 5 dpi na infectie. Schaalbalk: 50 μm. n = 4 muizen per groep. d IF-kleuring van eMyHC (rood) en Laminin (groen) werd uitgevoerd op de bovenstaande TA-spieren. Schaalbalk: 50 μm. e Het aantal eMyHC+-vezels per weergave werd gekwantificeerd. n = 4 muizen per groep. Van links naar rechts, p = 0.20, 0.00081, 0.052 en 0.012. f Links: IF-kleuring van Pax7 (rood) en Laminin (groen) werd uitgevoerd op de bovenstaande TA-spieren. Schaalbalk: 50 μm. Rechts: het aantal Pax7+ cellen per weergave werd gekwantificeerd. n = 4 muizen per groep. Van links naar rechts, p = 0.33, 0.041, 0.67 en 0.0052. g Links: IF-kleuring van Myod (rood) en Laminin (groen) werd uitgevoerd op de bovenstaande TA-spieren. Schaalbalk: 50 μm. Rechts: het aantal Myod+ cellen per weergave werd gekwantificeerd. n = 4 muizen per groep. Van links naar rechts, p = 0.0053, 0.0058, 0.12 en 0.020. Alle staafdiagrammen worden weergegeven als gemiddelde ± SD. Studenten t test (tweezijdig ongepaard) werd gebruikt om de statistische significantie te berekenen (b, e-g): *p < 0.05, **p < 0.01, ***p < 0.001. ns geen betekenis. Brongegevens worden geleverd als een brongegevensbestand.
H2B-verlies verhoogt DNA-schade en senescentie Atf3 iKO SC's
Om de consequentie van H2B-verlies in SC's verder op te helderen, onderzochten we de instabiliteit van het genoom en cellulaire veroudering, aangezien histon-eiwitten belangrijk zijn voor de integriteit van chromatine en de stabiliteit van het genoom.57,59. FISC's van iKO- of Ctrl-spieren werden behandeld met verschillende doseringen (0, 1000 of 2000J) UV-licht en er werd een komeettest uitgevoerd. Er werd inderdaad een aanzienlijke toename van de lengte van de komeetstaart waargenomen bij alle drie de doseringen in de iKO in vergelijking met de Ctrl (Fig. 9a), wat suggereert dat H2B-afname genomische instabiliteit veroorzaakt. Dit werd verder bevestigd door een veel hoger percentage γH2AX+-cellen in de gekweekte iKO-cellen vergeleken met de Ctrl (Fig. 9b, c). Om te onderzoeken of de genomische instabiliteit leidt tot cellulaire senescentie, werden cellen gedurende 9 dagen in groeimedium gekweekt en werd β-Gal-kleuring uitgevoerd; een toename van 19% van β-Gal+ cellen werd gedetecteerd in de iKO in vergelijking met de Ctrl (Fig. 9d). Consequent, een aanzienlijke opregulering van p16, p21 en p53 mRNA's (Fig. 9e) en een hoger niveau van p21- en p53-eiwitten werden ook gedetecteerd in de iKO (Fig. 9f). Al met al tonen de bovenstaande bevindingen aan dat H2B-verlies leidt tot verhoogde genoominstabiliteit en cellulaire senescentie in de Atf3 iKO-cellen. Verwacht werd dat overexpressie van H2B in de iKO-cellen de accumulatie van DNA-schade verminderde en cellulaire senescentie remde, zoals beoordeeld door verlaagd p21-eiwit (Fig. 9g) En p16, p21 en p53 mRNA's (Fig. 9h). Deze resultaten geven aan dat de verminderde H2B-expressie ervoor zorgt dat SC's replicatieve veroudering ondergaan.
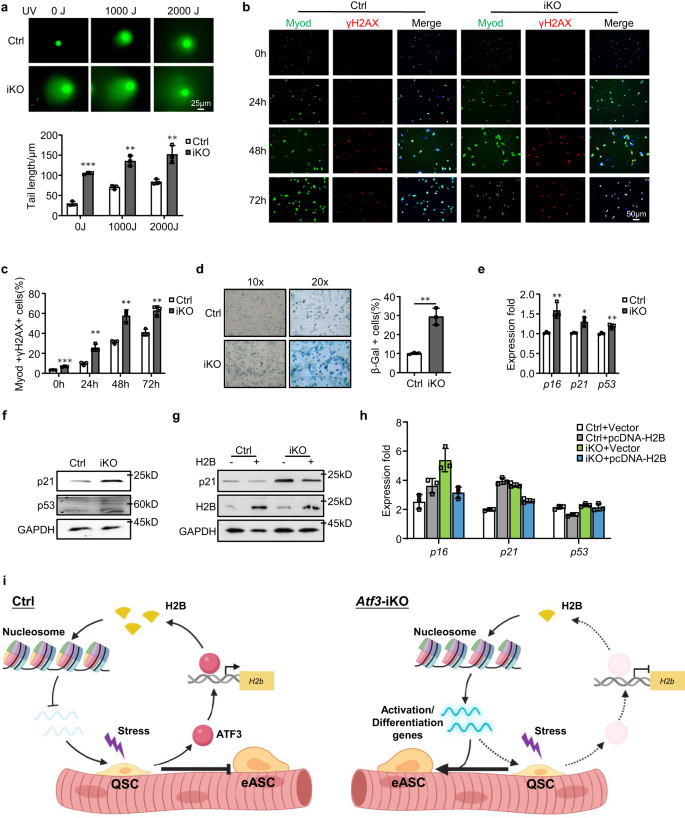
a Comet-assay werd uitgevoerd op Ctrl- of iKO SC's na 0, 1000 of 2000 J UV-behandeling. Schaalbalk: 25 μm. n = 3 muizen per groep. p = 0.000029, 0.0012 en 0.0071. b IF-kleuring van γH2AX (rood) en Myod (groen) werd uitgevoerd op SC's van Ctrl- of iKO-muizen na 0, 24, 48 of 72 uur gekweekt te hebben. Schaalbalk: 50 μm. c Kwantificering van de percentages Myod+/γH2AX+ cellen; n = 3 muizen per groep. p = 0.00014, 0.0024, 0.0021 en 0.0031. d β-Gal (blauwe) kleuring werd uitgevoerd op SC's van Ctrl- of iKO-muizen na 9 dagen kweken. n = 3 muizen per groep. p = 0.0017. e Expressie van de geselecteerde senescentiemarkergenen, waaronder p16, p21 en p53 in SC's van Ctrl en iKO werd gekwantificeerd door RT-qPCR. p = 0.0078, 0.011 en 0.0061. f p21- en p53-eiwitniveaus werden gedetecteerd door Western-blotting in SC's van Ctrl- en iKO-muizen. GAPDH werd gebruikt als laadcontrole. g, h SC's van Ctrl- en iKO-muizen werden getransfecteerd met een H2B tot overexpressie brengend of vectorcontroleplasmide. 96 uur na transfectie werd expressie van de aangegeven genen gedetecteerd door Western-blotting en RT-qPCR. GAPDH werd gebruikt als laadcontrole. i Schematisch model dat de functionele rol van ATF3 weergeeft bij het voorkomen van vroegtijdige activering van SC. In homeostatische spieren wordt de snelle inductie van ATF3 bij lichte stress bevorderd H2b expressie om de juiste positionering van het nucleosoom te behouden en de expressie van activeringsgenen te onderdrukken, waardoor vroegtijdige activering van SC's wordt voorkomen. Bij ATF3-verlies treedt H2B-deficiëntie op, wat leidt tot een veranderde positionering van het nucleosoom en opwaartse regulatie van activeringsgenen, waardoor SC's de rust doorbreken en het vroege activeringsstadium ingaan. Gemaakt met BioRender.com. Alle staafdiagrammen worden weergegeven als gemiddelde ± SD. Studenten t test (tweezijdig ongepaard) werd gebruikt om de statistische significantie te berekenen (a, c-e): *p < 0.05, **p < 0.01, ***p < 0.001. ns geen betekenis. Brongegevens worden geleverd als een brongegevensbestand.
Screening van andere functionele AP-1-familieleden in SC's en regeneratie
Na de bovenstaande holistische karakterisering van de functie en het mechanisme van ATF3 in SC's, probeerden we het onderzoek uit te breiden door te vragen of andere TF's uit de AP-1-familie, zoals ATF4, FOS, FOSB en JUNB, ook functionele rollen spelen in SC's, gezien hun vergelijkbare snelle en voorbijgaande inductiedynamiek tijdens SC vroege activering (Fig. 1b-d en S1c). Om het idee te testen, hebben we een functionele screening uitgevoerd met behulp van ons recent ontwikkelde Pax 7Cas9/AAV-sgRNA gemedieerd in vivo genoombewerkingsplatform64, die is gebaseerd op een Cre-afhankelijke Cas9 knock-in muizen en AAV-gemedieerde sgRNAs-afgifte. In het kort, één paar sgRNA's gericht op elke TF werd geselecteerd en verpakt in AAV9-virusdeeltjes; 4 × 1011 virale genomen (vg)/muis van AAV9-sgRNA werd intramusculair (IM) geïnjecteerd in de TA-spieren van Pax 7Cas9 muizen op postnatale (P) leeftijd van 10 dagen (P10). Voor de controlegroep (Ctrl) werd dezelfde dosis AAV9-virus met pAAV9-sgRNA-vectorruggengraat zonder enige sgRNA-insertie geïnjecteerd. De muizen werden vervolgens na acht weken opgeofferd voor SC-isolatie en analyse (Fig. S8a). Succesvolle DNA-bewerkingsefficiëntie werd bevestigd voor alle vier de TF's (aanvullend Fig. 8b-e); consequent werd knock-down (KD) van elk eiwit in verschillende mate (52.4, 83.6% - XNUMX, XNUMX%) gedetecteerd door WB (aanvullend in Fig. 7b-e) of IF (aanvullend Fig. 8f) ondanks ongewijzigde mRNA-niveaus (aanvullend Fig. 8g).
Om het effect van de bovenstaande AP-1 TF KD op SC-activering te onderzoeken, werden FISC's van elke mutant gekweekt voor in vitro EdU-assay (aanvullend Fig. 8a). Gelijkwaardig aan Atf3 KO, de Fos-KD-cellen vertoonden versnelde SC-activering terwijl de JunB-KD vertoonde afgestompte activering; de Atf4 or FosB-KD had daarentegen geen enkele impact (aanvullend Fig. 8h). Bij het meten van Pax7+-cellen op de ongeveer 2 maanden oude niet-verwonde spieren, werd interessant genoeg een verminderd aantal waargenomen op de Atf4, Fos or juniB-KD (aanvullend Fig. 8i), wat wijst op een verlies van SC-pool. Sinds de Atf4 or juniB-KD leidde niet tot vroegrijpe SC-activering, het verlies van het SC-nummer kan worden veroorzaakt door andere redenen, aangezien de deletie zeer vroeg in de postnatale fase plaatsvond64. De FosB-KD had geen invloed op de Pax7+ SC-pool.
Om de impact op het spierregeneratieve vermogen verder te onderzoeken, heeft BaCl2 werd 7 weken na de AAV-injectie geïnjecteerd (aanvullend Fig. 8a) en de regeneratie werd beoordeeld door H&E (aanvullend Fig. 8j), eMyHC (aanvullend Fig. 8k), Pax7 (aanvullend Fig. 8l) en MyoD (aanvullend Fig. 8m) kleuring op 5 dpi. Een verminderde regeneratie werd onthuld in de Atf4 en Fos-KD vermoedelijk vanwege de uitputting van SC's na langdurige verwijdering; De regeneratie in de JunB-KD was ook gecompromitteerd, waarschijnlijk als gevolg van een verminderde SC-pool; consistent werd geen invloed op de regeneratie waargenomen in de FosB-KD. Al met al blijkt uit onze resultaten dat deze AP-1-familieleden verschillende functies kunnen spelen bij SC-activering en spierregeneratie.
- Door SEO aangedreven content en PR-distributie. Word vandaag nog versterkt.
- PlatoData.Network Verticale generatieve AI. Versterk jezelf. Toegang hier.
- PlatoAiStream. Web3-intelligentie. Kennis versterkt. Toegang hier.
- PlatoESG. Automotive / EV's, carbon, CleanTech, Energie, Milieu, Zonne, Afvalbeheer. Toegang hier.
- Plato Gezondheid. Intelligentie op het gebied van biotech en klinische proeven. Toegang hier.
- ChartPrime. Verhoog uw handelsspel met ChartPrime. Toegang hier.
- BlockOffsets. Eigendom voor milieucompensatie moderniseren. Toegang hier.
- Bron: https://www.nature.com/articles/s41467-023-40465-w

