De L. album is een bekende bron van lignanen. Er zijn verschillende onderzoeken uitgevoerd naar hun fysiologische en biochemische eigenschappen en expressie van enkele sleutelgenen die betrokken zijn bij de biosynthese van lignanen in vitro kweektechnieken3,9,21. Deze studie was gericht op het herkennen van een gemeenschappelijke basis van transcriptieveranderingen, met name veranderingen geassocieerd met PTOX-biosynthese die optreden in verschillende organen tijdens in vivo culturen met meerdere stress.
De expressieprofielen vertoonden duidelijk verschillen in het aantal opwaarts en neerwaarts gereguleerde DEG's onder meerdere spanningen. Er werden minder DEG's waargenomen in de spanningen van deze plant met betrekking tot de wortels (zoals Al-toxiciteit) in vergelijking met de scheuten. Deze verschillen worden gedreven door verschillen tussen datasets die verschillende bronnen kunnen hebben, zoals biologische of technische variabiliteit.
Identificatie van een groot aantal DEG's onder droogtestress gaf aan dat een aanzienlijk deel van het transcriptoom door deze stress was bezet22. Onder droogtestress ontstaan de verschillen tussen tolerantie en vatbare genotypen in genexpressiepatronen en fenotypes10. Ook is waargenomen dat het expressieniveau van responsieve stressgenen gewoonlijk hoger is bij tolerantiegenotypes13. Daarom bevestigen deze resultaten het concept dat vlas een betere droogtetolerantie heeft dan andere stressfactoren10.
Volgens de geïdentificeerde DEG's werden 20 algemeen gereguleerde genen die reageren op biotische en abiotische stress en 38 algemeen gereguleerde genen die reageren op abiotische stress overlapt met behulp van Venn-diagramanalyses in respectievelijk de scheut en de wortel. Dan slechts één algemeen gereguleerd gen, endochitinase EP3, werd geïdentificeerd met een significante toename van beide organen. De EP3katalyseert de hydrolyse van chitine, is betrokken bij de ontwikkelingsprocessen van planten, het genereren van signaalmoleculen, afweerreacties van planten en geprogrammeerde celdood23,24. De uitdrukking van de EP3 gen verhoogd als reactie op droogte, zoutgehalte, behandelingen met UV-licht, exogene elicitorbehandeling, verwonding en aanval door pathogenen11,23,24. Bovendien nam het β-1,3-glucanase-gen toe in resultaten met betrekking tot de scheut, waarvan eerder was aangetoond dat het cruciaal was voor vlasresistentie tegen Fusarium spp.25. Vlasplanten met overexpressie van β-1,3-glucanase, het genereren van pectinaseremmers en het gebruik van chitinasen en peroxidasen verhoogden de productie van SM's en veranderde celwandsamenstellingen, wat leidde tot de constructie van een barrière voor schimmelgroei26.
Biologische processen, waaronder secundair metabolisme, stress, ontwikkeling, celwand, vetmetabolisme en eiwitafbraak, werden gewoonlijk veranderd door alle stress. Een dergelijke gemeenschappelijke regulering weerspiegelt de flexibiliteit van biologische systemen door de aanpassing van complexe metabolismenetwerken als reactie op stimulerende middelen tijdens de evolutie van het onbeweeglijke leven van de plant.11.
De op stress reagerende gemeenschappelijke GO-termen bestonden uit AP2 / ERF-, HB-7- en NAC-transcriptiefactorfamilies die aanzienlijk werden opgereguleerd in ABA-behandeling en droogtestress en gedownreguleerd in Fusarium behandeling en K+ tekort. De studie van individuele stress heeft 11 top-TF's geïdentificeerd, waaronder bHLH (Basic helix-loop-helix), C2H2, NAC, MYB, ERF, bZIP, WRKY, MYB, DREB, HSF en NFYA10 als bekende belangrijkste regulatoren van abiotische resistentieroutes onder herhaalde droogte in vlas10. NAC TF's droegen ook bij aan de stressreactie op maïs, rijst en vlastolerantie voor aluminiumstress. Deze TF's, samen met MADS-box, passen de groei en ontwikkeling van planten aan en brengen veranderingen in de celwand met zich mee, wat leidt tot tolerantie voor aluminium13. Van de meeste TF's die verband houden met kaliumgebrek is aangetoond dat ze behoren tot MYB, bHLH, NAC, B3, bZIP, WRKY en AP2/ERF, die deelnemen aan fysiologische plantprocessen, stressbestendigheid en secundair metabolisme18. De stresssignaleringsroutes delen gemeenschappelijke componenten, waaronder ROS, calciumionen, hormonen, TF's en mitogeen-geactiveerde proteïnekinase (MAPK) cascades27.
De root GO-termen toonden aan dat biologische processen, namelijk TCA/org-transformatie, oxidoreductase-activiteit, celwand, transport, hormoonmetabolisme en receptorkinasesignalering, specifiek worden gemoduleerd onder abiotische stress. In een onderzoek naar vlas onder individuele stress, waren GO-termen van oxidoreductase-activiteit, met name peroxidasen, celwand, ionenhomeostase en stressrespons het meest veranderd onder ongunstige omstandigheden van de pH en het Zn-tekort12.
Sommige van onze resultaten kwamen overeen met de afzonderlijke onderzoeken, andere waren er tegen. Deze individuele stressfactoren kunnen verschillende aanpassingsreacties veroorzaken, die verschillende of gedeelde componenten in planten bevatten. Wanneer planten tegelijkertijd stresscombinaties tegenkomen, kan dit vergelijkbare of tegengestelde moleculaire, fysiologische en metabolische reacties vereisen. De precieze keuze van welke aanpassingsstrategie tijdens meervoudige stress vermoedelijk zal worden beïnvloed door factoren zoals de intensiteit van elke individuele stress, het tijdsverloop van stress, plantleeftijd en genotype (tolerant of vatbaar voor een van de individuele stress)28. Het type aanpassing kan de accumulatie van SM's beïnvloeden, waardoor ruimtelijke en temporele modulatie van de biosynthetische routes wordt veroorzaakt, waardoor de overlevingskans in langdurige stressvolle omgevingen wordt verbeterd29.
MapMan-analyse categoriseerde SM's in 16 groepen. Fenylpropanoïde- en flavonoïderoutes werden sterk beïnvloed door alle spanningen. De hoogste overvloed aan transcripten was gerelateerd aan flavonoïde- en fenylpropanoïde-routes onder droogtestress. De toename van deze verbindingen was extreem gerelateerd aan de balans van koolhydraten tussen bronnen en putten. Er is ook gemeld dat het waterpotentieel in de plant afnam onder ernstige droogtestress, wat leidde tot het transport van oplosbare suikers. Daardoor nam de ophoping van flavonoïden en fenolen toe30,31. Uitkomsten met betrekking tot droogtestress in dit werk toonden een verminderde vijfvoudige (-5) transcriptovervloed in de glucosinolaatroute. Volgens deze resultaten verminderde droogtestress het glucosinolaatgehalte aanzienlijk Boechera holboellii Hoorn. Á.Löve & D.Löve en nog wat Brassica carinata A.Braun cultivars, terwijl sommige B. karinata cultivars vertoonden een significante toename van glucosinolaat32,33. Behandeling met kaliumsulfaat verminderde het glucosinolaatgehalte tijdens droogte op canola in vergelijking met de onbehandelde planten34. Roots vertoonde enkele veranderingen in de overvloed aan transcripties van SM's. Op stress reagerende transcripten worden echter meestal gewijzigd onder ongunstige omstandigheden. SM's, zoals sinapinezuur, lignine en flavanolen, met defensieve rollen, namen toe Sinopodophyllum hexandrum Royle onder water tekort35.
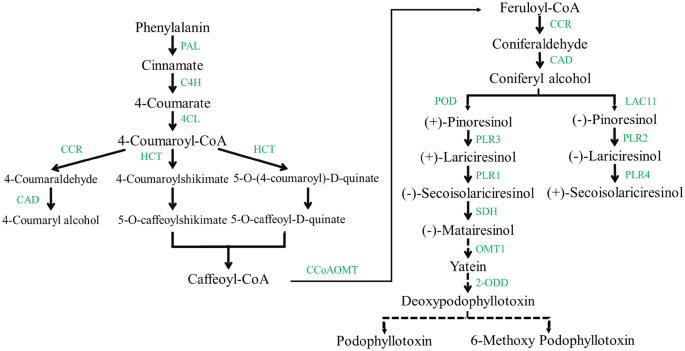
Verschillende fenolverbindingen, waaronder flavonoïden, monolignolen, lignanen, ligninen, coumarinen en hydrolyseerbare tanninen, worden gevormd via de fenylpropanoïde route36. De Linum geslacht, in het bijzonder L. album, bevat de hoogste niveaus van lignanen, vooral PTOX. Veel onderzoeken hebben echter de lignan-biosyntheseroute onderzocht, de lignaanroute totdat het eindproduct PTOX niet volledig is opgehelderd.8,20.
Deze route begint met de deaminering van fenylalanine en synthetiseert 4-coumaroyl-CoA door 4CL. Vervolgens wordt 4-coumaroyl-CoA via verschillende reacties door HCT omgezet in caffeoyl-CoA, dat twee verschillende stappen katalyseert, gevolgd door methylering via CCoAOMT en feruloyl-CoA synthetiseert1,37. Feruloyl-CoA wordt omgezet in coniferylalcohol door twee reductiereacties door CCR en CAD8,38. Aangezien coniferylalcohol bekend staat als een kritische voorloper in de biosynthese van PTOX, werden de bovengenoemde stappen als stroomopwaarts beschouwd39. In de huidige studie bevat CAD het grootste aantal transcripten in de stroomopwaartse stappen die ruimtelijke expressiepatronen laten zien en mogelijk verschillende functionele rollen hebben in specifieke organen. Bijvoorbeeld, OsCCR10 gaat in reactie op droogte in rijstwortel40. Knock-out van OsCCR10 met het CRISPR/Cas9-systeem onthulde dat droogtetolerantie rijst verminderde door een afname van het ligninegehalte in de wortel41. Na coniferylalcohol werden de later gespecificeerde stappen van PTOX en de daarvan afgeleide biosynthese als stroomafwaarts beschouwd. Deze stappen beginnen met het koppelen van twee moleculen coniferylalcohol om pinoresinol-enantiomeren te krijgen door een oxidase (LAC11) of peroxidase (POD) met behulp van dirigent-eiwitten, afhankelijk van de plantensoort42,43,44. L. usitatissimum genereert zowel enantiomeren (-)- als (+)-pinoresinol, gevolgd door de stapsgewijze reductieve omzetting in lariciresinol en vervolgens SECO via PLR (1–4)45. (-) SECO, resulterend uit (+)-pinoresinol, wordt gekatalyseerd door de werking van SDH tot matairesinol. Vervolgens wordt matairesinol door verschillende enzymatische reacties omgezet in deoxypodofyllotoxine P. hexaandrum, zoals PhOMT3, CYP71CU1, PhOMT1 en 2-ODD. De genen die coderen voor enzymen die geassocieerd zijn met stappen tussen matairesinol en deoxypodofyllotoxine zijn echter nog niet geïdentificeerd in Linum8,46,47. Deze studie realiseerde enkele van deze genen. De transcriptomische analyse van L. usitatissimum onthulde dat de hoogste expressieniveaus van LAC11, POD, 4CL en SDH genen stonden onder droogtestress. De wortels onderworpen aan abiotische stress toonden de neerwaartse regulatie van het expressieniveau van deze enzymen aan. Omgekeerd, kwantitatieve uitdrukking van SDH het gebruik van qRT-PCR liet een stijgende trend zien in L. album wortels en verschilde in zijn scheuten onder alle behandelingen. Verder, 2-ONEVEN, exclusief droogtestress, en OMT1 vertoonde een stijgende lijn in L. album onder alle stress, in overeenstemming met eerder gerapporteerde studies in P. hexaandrum. Deze route evolueerde echter onafhankelijk in de twee soorten20. Kwantitatieve expressie van genen gerelateerd aan PTOX-biosynthese, bestudeerd in verschillende organen van P. hexaandrum, toonde dat aan SDH, CADXPERT / LANDXPERT, CCR, en cinnamaat 4-hydroxylase genen namen meer toe in wortelstokken dan in wortels37. De verhoogde genexpressies gerelateerd aan groei en ontwikkeling en PTOX-biosynthese werden ook gemeld bij 15 °C in S. hexaandrum. Terwijl de genexpressie en het PTOX-gehalte afnam en de genen die reageerden op stress domineerden bij 25 °C in deze plant1. In het onderzoek van Kumari et al. (2022), genen van fenylalanine ammoniak-lyase (ShPAL), Sh4CL, ShC3H, ShCCoAOMT, ShCOMT, ShCAD, ShDPO, ShPLR en ShSDH opgereguleerd, evenals een verhoogd PTOX-gehalte in de wortel onder droogtestress. Er was echter geen bewijs van PTOX in het blad35.
High-performance liquid chromatography (HPLC)-analyse, consistent met qRT-PCR-resultaten, toonde een toename van het SECO-gehalte en opregulatie van drie geselecteerde genen aan (SDH, OMT1 en 2-ONEVEN) leidde tot de productie van de hoogste PTOX in de wortels na een combinatiebehandeling van 48 uur. Tegelijkertijd produceerden scheuten PTOX door de voorloper van SECO te consumeren. Onder droogtestress in de wortel om 12 uur een overvloedige hoeveelheid SECO en overexpressie van 2-ONEVEN veroorzaakte de omzetting van 6-MPTOX en PTOX tijdens stapsgewijze reacties1. Het PTOX-gehalte daalde echter vanwege de neerwaartse regulering van 2-ONEVEN na 48 uur, volgens qRT-PCR. Andere enzymen die verband houden met de biosynthese van lignan zijn waarschijnlijk betrokken bij de 6-MPTOX-generatie, dus PTOX nam af onder droogtestress. Ondanks het hoge SECO-bedrag en de overexpressie van 2-ONEVEN (in de shoot na 48 uur), nam PTOX aanzienlijk af vanwege de neerwaartse regulatie van SDH en OMT1. de K+ tekort en droogtestress hadden vaak tegengestelde accumulatiepatronen van drie geselecteerde lignanen. De verschillende reacties op individuele en gecombineerde spanningen in L. album wordt het complexe regulerende mechanisme van de biosynthese van lignanen gesuggereerd, dat verder onderzoek vereist. De uitdrukking van PLR in S. hexaandrum is in tegenspraak met het PTOX-gehalte in de verschillende organen, wat suggereert dat het PTOX-producerende weefsel niet noodzakelijkerwijs de pool is48.
Toepassing van abiotische en biotische elicitors, waaronder chitosan, methyljasmonaat, salicylzuur, gistextract en Ag+, heeft grotendeels vastgesteld dat de expressie van genen geassocieerd met lignaanbiosynthese en lignaangehalte verbeterd in Linum spp. in vitro49,50,51,52. Ook een studie uitgevoerd op verschillende toetredingen van L. album onder droogtestress vertoonden verschillende patronen op basis van fysiologische en biochemische reacties53.
Over het algemeen zijn de resultaten van qRT-PCR en HPLC-analyse met transcriptomische analyse consistent, wat eerdere studies bevestigt. Hoewel ze in sommige gevallen kunnen verschillen vanwege verschillen in soort of genotype, type cultuur en stressintensiteit. de K+ Deficiëntie en droogtestress hadden vaak tegenovergestelde opwaartse/neerwaartse regulatiepatronen, waarbij de negatieve synergetische effecten veel kosten voor de plant met zich meebrachten. Hoewel het opwaartse/neerwaartse regulatiepatroon van fusarium vergelijkbaar was met dat van K+ tekort, was droogtestress vergelijkbaar met ABA-behandeling.
Daarom zouden strategieën op basis van transcriptoom voor soorten die waarschijnlijk lignanen accumuleren, helpen bij het identificeren van gemeenschappelijke kenmerken tussen soorten en omgevingssignalen om de PTOX-biosyntheseroute te verduidelijken.
- Door SEO aangedreven content en PR-distributie. Word vandaag nog versterkt.
- PlatoAiStream. Web3 gegevensintelligentie. Kennis versterkt. Toegang hier.
- De toekomst slaan met Adryenn Ashley. Toegang hier.
- Koop en verkoop aandelen in PRE-IPO-bedrijven met PREIPO®. Toegang hier.
- Bron: https://www.nature.com/articles/s41598-023-36102-7

